
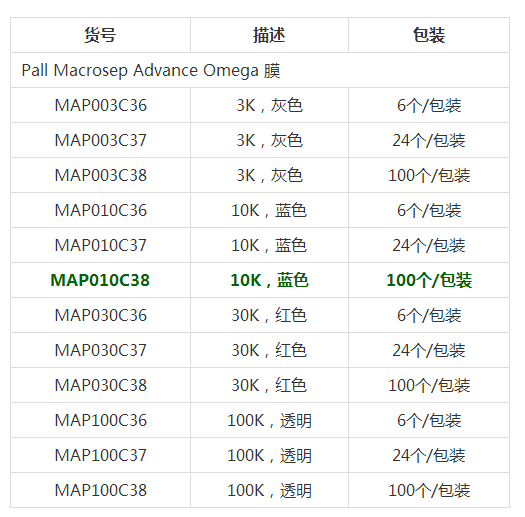
- 型号 MAP010C38
- 品牌 PALL颇尔
- 【简单介绍】
PALL颇尔 离心浓缩管 MAP010C38
· 可迅速把20ml样品浓缩到0.5ml。· 回收率高,通常可达90%以上。
· Pall Omega膜和聚丙烯外壳的蛋白质结合率低,能把由非特异性结合引起的损失降低到zui低。
· 多功能的Pall Omega膜可用于各种MWCO中。
· 以颜色编码,易于鉴别。
PALL颇尔 离心浓缩管 MAP010C38
说明
· 可迅速把20ml样品浓缩到0.5ml。
· 回收率高,通常可达90%以上。
· Pall Omega膜和聚丙烯外壳的蛋白质结合率低,能把由非特异性结合引起的损失降低到zui低。
· 多功能的Pall Omega膜可用于各种MWCO中。
· 以颜色编码,易于鉴别。
Pall 颇尔 Macrosep Advance离心浓缩管
应用
· 对蛋白样品进行浓缩和脱盐
· 对层析后的样品或梯度洗脱后的样品进行缓冲液的置换和脱盐
· 从细胞培养上清液或者裂解液中回收生物分子
· 从水溶液中去除颗粒
技术规格
制造材料
滤才: Omega (低蛋白吸附改性的聚醚砜) 和Supor膜(polyethersulfone) 样品池, 滤液接收器, 和盖子:聚丙烯
液板: 聚乙烯
有效过滤面积:7.2 cm2
尺寸:直径:50 mm (1.9 in.),长度:12.0 cm (4.7 in.)
容量:zui大样品体积:20 mL,zui终浓缩体积:根据使用的转头而定,zui少可达450?L。滤液槽体积:22 mL,残留体积(膜/支撑物):80 ?L
操作温度范围:0 – 40 °C (32 – 104 °F)
pH范围:1-14
zui大离心力:5,000 x g (超滤),14,000 x g (微滤)
离心:适合使用标准的50 mL的管子
消毒情况:未灭菌。在使用前用70%的乙醇进行过滤,已进行消毒。性能
转头的选择决定zui终的浓缩液体积
转角 残留体积
吊桶式转头 450?L
45°固定角 1.2-1.5mL
34°固定角 1.5 mL
Pall 颇尔 Macrosep Advance离心浓缩管
PALL颇尔 离心浓缩管 MAP010C38
标签归档:amp
FSD™ Fluor 荧光染料 Fluorescent FSD™ Fluor Dyes
- 产品特性
- 相关资料
- Q&A
- 参考文献

FSD™ Fluor 荧光染料
Fluorescent FSD™ Fluor Dyes
BioActs 品牌介绍
AdipoGen Life Sciences 旗下的子品牌 BioActs 研发并生产高灵敏度、高纯度、高质量的新型和传统生物荧光探针,如荧光染料、体外诊断产品用光学材料、近红外 (NIR) 材料和体内研究用造影剂。BioActs 的产品已被世界各地的大学、研究所和企业作为光学材料和探针使用,广泛用于医学、制药和化学研究领域的基础和应用研究。
BioActs 专注于提供多种生化标记染料(含 Flamma® Fluor 染料或 FSD Fluor™ 染料)、细胞分析染料(如 ApoFlamma®、CytoFlamma® 或 MitoFlamma®)、体内成像染料(如 NpFlamma®、AngioFlamma ®、NIR 染料或生物发光染料)或偶联二抗。此外,BioActs 可提供多种核酸标记染料、纳米粒子和磁珠,并可提供全方位的定制标记服务。

新型 FSD™ Fluor 荧光染料
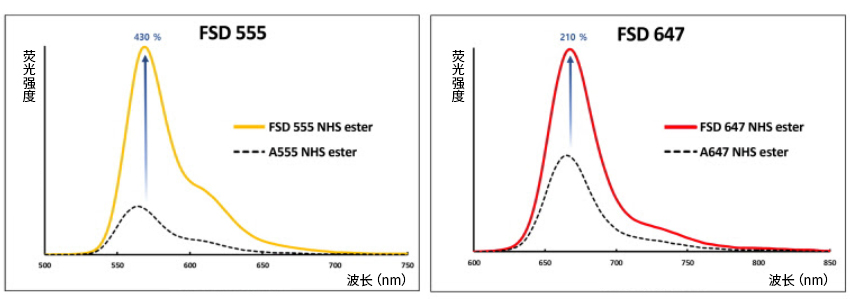
FSD™ Fluor 系列是新型的荧光染料,与传统荧光染料相比,具有出色的荧光强度和高量子产率。与生物分子靶标(抗体、肽、蛋白质等)结合后,其荧光强度仍相当出色。FSD™ Fluor 系列具有多种用途,适用于大多数荧光设备和广泛的应用研究。另有多种反应官能团可供选择。
– 卓越的荧光性能
– 高量子产率
– 多种反应官能团可供选择
● 染料荧光强度的比较
● 染料-抗体偶联物的荧光强度比较

● 各染料适用波长一览
● 染料的相对量子产率的比较

※ 本页面产品仅供研究用,研究以外不可使用。
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| BCT-RSA4445-MC05 | Rabbit anti-goat IgG, FSD 488 兔抗山羊IgG抗体, FSD 488 |
0.5 mg | |
| BCT-RSA4445-M001 | Rabbit anti-goat IgG, FSD 488 兔抗山羊IgG抗体, FSD 488 |
1 mg | |
| BCT-RSA4455-MC05 | Rabbit anti-goat IgG, FSD 555 兔抗山羊IgG抗体, FSD 555 |
0.5 mg | |
| BCT-RSA4455-M001 | Rabbit anti-goat IgG, FSD 555 兔抗山羊IgG抗体, FSD 555 |
1 mg | |
| BCT-RSA4495-MC05 | Rabbit anti-goat IgG, FSD 594 兔抗山羊IgG抗体, FSD 594 |
0.5 mg | |
| BCT-RSA4495-M001 | Rabbit anti-goat IgG, FSD 594 兔抗山羊IgG抗体, FSD 594 |
1 mg | |
| BCT-RSA4465-MC05 | Rabbit anti-goat IgG, FSD 647 兔抗山羊IgG抗体, FSD 647 |
0.5 mg | |
| BCT-RSA4465-M001 | Rabbit anti-goat IgG, FSD 647 兔抗山羊IgG抗体, FSD 647 |
1 mg | |
| BCT-RSA4405-MC05 | Rabbit anti-goat IgG, FSD 750 兔抗山羊IgG抗体, FSD 750 |
0.5 mg | |
| BCT-RSA4405-M001 | Rabbit anti-goat IgG, FSD 750 兔抗山羊IgG抗体, FSD 750 |
1 mg | |
| BCT-RSA4485-MC05 | Rabbit anti-goat IgG, FSD 800 兔抗山羊IgG抗体, FSD 800 |
0.5 mg | |
| BCT-RSA4485-M001 | Rabbit anti-goat IgG, FSD 800 兔抗山羊IgG抗体, FSD 800 |
1 mg |
ClearSee™ 植物科学新技术 植物透明化试剂
- 产品特性
- 相关资料
- Q&A
- 参考文献
植物科学新技术 植物透明化试剂
ClearSee™
ClearSee™ 是一种仅需固定、洗净、透明化三步简单操作即可实现植物组织透明化的试剂。作为植物科学的研究工具,期待可以阐明细胞水平的现象和整个个体之间的系统联系。
HP数据:由名古屋大学大学院理学研究科 栗原大辅特任助教、水多阳子研究员提供

◆操作步骤
使用 ClearSee™ 的透明化方法,只需1)固定、2)洗净、3)透明化三个步骤。详细方法可参考随产品附赠的使用说明书。
以下,是拟南芥叶和幼株的使用案例。
1. 固定
① 使用4%甲醛溶液固定样本*1。

② 将样本放入干燥机中,用真空泵减压,在室温下静置30分钟,并固定。
② 以上操作重复两次。
② 为了防止植物样品在减压过程中飞出,建议打开管盖,用封口膜密封,并用针在上面扎孔*2,3。

2. 洗净
去除固定液,加入1 mL的1×PBS缓冲液,静置1分钟。然后去除1×PBS缓冲液,重新加入1 mL的1×PBS缓冲液,静置1分钟。
以上操作重复两次。
3. 透明化处理
去除1×PBS缓冲液,添加1.3 mL的ClearSee™。
打开管盖后用封口膜密封,并用针在上面扎孔。放入干燥机中,用真空泵减压,并在室温下静置1小时。
取下封口膜,盖好管盖。避光,室温静置,透明化*4。
如有需要可更换新的ClearSee™。
4. 透明化过程

图4. ClearSee™中的拟南芥叶和幼株

图5. 可以观察到样本周围叶绿素(1-2天后)

图6. 更换新的 ClearSee™ 后的样本

图7. 透明化的拟南芥叶和动株(3-5天后)
使用ClearSee™ 进行透明化需要的物品
|
1. 高压灭菌锅
2. 局部排气系统(通风室等) 3. 真空泵 4. 干燥机 5. 60°C 恒温器 6. 荧光显微镜 7. 锥形管 |
8. 微量离心管 9. 封口膜 10. 针 11. 镊子 12. 盖玻片 13. 载玻片 14. 凡士林油 |
注意
※ 由于植物活体样本含有空气,添加溶液时植物就会漂浮起来。请尽可能地将植物样本浸入固定液中。
※ 请将干燥机设定在-90 kPa左右。根据植物样本的不同,有时减压会过强。减压过强的话,会破坏植物组织和细胞。开始减压后,由于会抽出植
※ 物样本中的空气,会产生气泡。为了让气泡消失,请缓慢调整减压的压强。
※ 从减压状态恢复至大气压强时,请缓慢进行。如果急速恢复至大气压强的话,会破坏植物组织和细胞。
※ 透明化的时间根据植物种类和组织而异,请参照以下的时间。
※ 根:1~2天、叶和幼株:4~7天、雌蕊:2周、成熟组织:4周。
◆观察案例
5. 观察
在载玻片上用凡士林或油脂制作一个围栏,放入适量 ClearSee™,用镊子将样本移入,盖好盖玻片,并用荧光显微镜观察。由于 ClearSee™干燥后会产生沉淀,所以用凡士林或油脂来防止其干燥。观察结束后,可以将植物样本放回装有 ClearSee™ 的微管中进行保存。
另外,用 ClearSee™ 透明化的样本亦可用荧光色素进行染色。细胞核染色时,向 ClearSee™ 中添加终浓度为 10 μg/ mL 的双苯甲酰亚胺 H33258 三盐酸盐,双苯甲亚胺 H33342 或 DAPI,细胞壁染色时,使用终浓度为 1 mg/mL,静置1小时后,更换新的 ClearSee™ ,再静置1小时后,在荧光显微镜下进行观察。

图8. 经透明化处理的拟南芥叶在明场中呈透明状态,叶绿素的红色自发荧光消失。
图8. 由于可以对整片叶进行观察,因此也可以观察到维管束内的荧光蛋白。比例尺为 200 μm。

图9. 使用 ClearSee™ 透明化的完整的雌蕊的荧光观察。
图2. 用4个颜色(蓝色,绿色,黄色,红色)的荧光蛋白标记花粉管。
图2. 根据参考文献改编转载。

图10. 使用 ClearSee™ 处理4天的拟南芥的根。
图10. 生长素应答细胞的细胞核用绿色、全身的细胞核用紫色显示。
图10. 根据参考文献改编转载。
■其他植物的透明化案例
使用 ClearSee™ 透明化6天后的蓝猪耳、矮牵牛、烟草、番茄、黄瓜、稻叶。
稻叶在透明化时,由于叶子的表面有蜡层,因此很难将溶液渗入组织里,通过在氯仿等有机溶剂中浸泡 10~30 秒除蜡后,开始固定的步骤。

图11. 使用 ClearSee™ 透明化的各种植物叶片(6天后)
◆参考文献
Kurihara, D. et al.: Development, 142, 4168-4179 (2015).
点击此处查看产品宣传页
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 031-25151 | ClearSee™ | 50 mL | 植物透明化用 |
| 160-16061 | 多聚甲醛 Paraformaldehyde |
100 g | 组织固定用 |
| 162-16065 | 多聚甲醛 Paraformaldehyde |
500 g | 组织固定用 |
| 167-25981 | 16w/v% 低聚甲醛溶液 16w/v% Paraformaldehyde Solution, Methanol free |
1 mL×10A | 电子显微镜用 |
| 163-25983 | 16w/v% 低聚甲醛溶液 16w/v% Paraformaldehyde Solution, Methanol free |
10 mL×10A | 电子显微镜用 |
MagCapture™ microRNA分离试剂盒系列 MagCapture™ microRNA Isolation Kit 系列
- 产品特性
- 相关资料
- Q&A
- 参考文献
 MagCapture™ microRNA 分离试剂盒系列
MagCapture™ microRNA 分离试剂盒系列
本公司提供可实现从 Ago 免疫沉淀到RNA纯化一系列更简便操作的「microRNA Isolation Kit」系列产品。通过使用该系列产品,可便利地从人或小鼠的细胞、组织中回收 Ago 蛋白质结合 microRNA 以及目标 mRNA。另外,从培养上清、血清、血浆中也可回收游离状态的 Ago 蛋白质结合 microRNA。回收后的 RNA 可用于定量PCR、微阵列芯片(Microarray)、新一代序列分析等各种应用。该系列产品扩充至有采用了磁珠的 MagCapture™ microRNA Isolation Kit 和采用硅珠的,共8种试剂盒。

◆MagCapture™ microRNA Isolation Kit(磁珠型)
该系列有抗人 Ago2 抗体(4G8)、抗鼠 Ago2 抗体(2D4)、抗 Ago1-4 抗体(1G3)包被磁珠的三种试剂盒以及含有可固定任意抗体的 Protein G 磁珠试剂盒,共计4款产品。这些试剂盒都采用磁珠,可更方便地进行重复性良好的免疫沉淀操作。在免疫沉淀后的纯化 RNA 步骤中,采用能获得高纯度 RNA 的离心柱法;此外,也可以采用含有抗 Ago 抗体包被磁珠的3种试剂盒的更简便的一步法。
试剂盒内容(例:Human Ago2, 10次用)
● Anti-Human Ago2 Magnetic Beads Solution——600 μL×1支
● Cell Lysis Solution——20 mL×1支
● Washing Solution for IP——40 mL×1支
● Elution Solution I for IP—— 500 μL×1支
● Elution Solution II for IP——500 μL×1支
● Binding Solution for Spin Column——2 mL×1支
● Binding Enhancer for Spin Column——100 μL×1支
● Washing Solution I for Spin Column——3 mL×1支
● Washing Solution II for Spin Column—— 4 mL×1支
● Elution Solution for Spin Column——1 mL×1支
● Spin Column/Collection Tube——10支
◆概述图

|
产品名称 |
使用抗体克隆号 |
适应物种 |
对应样本 |
回收RNA |
IP B/F 分离 |
RNA提纯 |
|
MagCapture™ microRNA Isolation Kit, Human Ago2 |
4G8 |
人、猴、狗 |
细胞/ 组织/血清/ 血浆 |
microRNA 和目标mRNA |
磁力分离 |
离心柱法、 一步法 |
|
MagCapture™ microRNA Isolation Kit, Mouse Ago2 |
2D4 |
小鼠、大鼠、狗 |
||||
|
MagCapture™ microRNA Isolation Kit, Ago 1-4 |
1G3 |
人、猴、狗, |
microRNA |
|||
|
MagCapture™ microRNA Isolation Kit, Protein G |
随意 |
参照抗体 |
细胞/组织 |
参照抗体 |
离心柱法 |
◆MagCapture™ microRNA Isolation Kit, Human Ago2 实验案例(人)
从人细胞系中回收 microRNA
用变性聚丙烯酰胺凝胶电泳法(银染色)确认通过本产品(MagCapture™ microRNA Isolation Kit, Human Ago2)从人K562细胞(1×107 cells)中获得的 microRNA(图1)。结果表明,使用本产品可获得高纯度的 microRNA。此外,通过定量 PCR 法(miR-92a 量),与使用原产品 microRNA Isolation Kit,Human Ago2 所获得的 microRNA 量相比较可知(图2),本产品的离心柱法显示与原产品几乎一样的 microRNA 回收性能;另外,在一步法中显示出了比原产品更优秀的 microRNA 回收性能。
 |
 |
| 图1. 银染色检测从人K562细胞纯化的 microRNA | 图2. 人K562细胞中纯化 microRNA 回收量的比较 |
|
LaneM:标记 Lane1:合成RNA寡聚核苷酸(23base)1ng Lane2:本品(Spin Column法) Lane3:本品(One Step法) |
从人血浆中回收结合游离 Ago 蛋白质的 microRNA
通过定量 PCR 法比较使用本产品和原产品从 200 μL 正常人混合血浆中获取的microRNA 量。结果显示,本产品的离心柱法与原产品具有几乎一样的microRNA回收性能;另外,在一步法中microRNA 回收量比原产品多。(见图3)

图3.人血浆中纯化 microRNA 回收量的比较
microRNA 的目标mRNA回收
向 miR-122 低表达的肝癌细胞株 HepG2 细胞(5×105 cells)中导入人工合成的 miR-122 和调节用荧光素酶 siRNA(Luc siRNA),使用本产品和原产品获得配制的 Ago2 IP RNA 以及用 ISOGEN® 配制的 total RNA。通过定量PCR法比较各RNA种所含 miR-122 的量(图4)和其目标mRNA(目标基因中 Aldo A mRNA 的量用 GAPDH mRNA 的量补充)。(图5)。结果表明,与原产品一样,通过本产品获得的目标 mRNA中,Aldo A mRNA 和 miR-122一同被浓缩,由此可知使用本产品可以获得 microRNA 的目标 mRNA。另一方面,使用 ISOGEN® 获得的 total RNA 中,Aldo A mRNA 随着 miR-122 的增加而逐渐分解减少。

图4.miR-122回收量的比较

图5.目标mRNA回收量的比较
◆MagCapture™ microRNA Isolation Kit, Mouse Ago2 实验案例(小鼠)
从小鼠细胞中回收microRNA
用变性聚丙烯酰胺凝胶电泳(银染色)确认通过使用本产品(MagCapture™ microRNA Isolation Kit, Mouse Ago2)从小鼠P388D1细胞(2×107 cells)中获得的 microRNA。(图1)结果表明,使用本产品可获得高纯度的microRNA。此外,通过定量PCR法(miR-92a量),与使用原产品(microRNA Isolation Kit, Human Ago2)所获得的microRNA量相比较可知(图2),本产品的离心柱法显示与原产品几乎一样的microRNA回收性能;另外,在一步法中显示出了比原产品更优秀的microRNA回收性能。
|
|
|
|
图1.银染色检测从小鼠P388D1细胞纯化的microRNA LaneM:标记 Lane1:合成RNA寡聚核苷酸(23base)1ng Lane2:本品(Spin Column法) Lane3:本品(One Step法) |
图2.小鼠P388D1细胞纯化microRNA回收量的比较 |


从大鼠血浆中回收游离Ago2蛋白质结合microRNA
通过定量PCR法(miR-92a量)比较使用本产品和原产品从 200 μL大鼠血浆中获取的microRNA量。结果表明,本产品的离心柱法与原产品具有几乎一样的microRNA回收性能;另外,在一步法中microRNA回收量比原产品大。(图3)

图3.人血浆中纯化microRNA回收量的比较
◆MagCapture™ microRNA Isolation Kit, Ago 1-4 实验案例(人)
回收Ago1-4结合microRNA
分别使用 Ago1, Ago2, Ago3, Ago4特异性抗体对HeLa细胞中各RNA片段免疫沉淀处理后,进行微阵列芯片(3D-Gene® ,东丽)分析。结果表明miR-22和miR-16多数被Ago2抗体吸收。另一方面,miR-1260b, miR-4286多数被Ago1, Ago3, Ago4吸收。(表1)
基于此解析结果,验证使用本产品(MagCapture™ microRNA Isolation Kit, Ago1-4),是否可回收原产品(microRNA Isolation Kit, Human Ago2)中被Ago1, Ago3, Ago4吸收较为困难的microRNA。
表1. 微阵列芯片分析结果
|
信号强度 |
信号比 |
||||||
|
Name |
Ago1 IP |
Ago2 IP |
Ago3 IP |
Ago4 IP |
Ago2/Ago1 |
Ago2/Ago3 |
Ago2/Ago4 |
|
hsa-miR-22 |
3521.4 |
21696.5 |
3771.2 |
292.4 |
6.16 |
5.75 |
74.20 |
|
hsa-miR-16 |
6886.3 |
28527.2 |
5013.4 |
582.5 |
4.14 |
5.69 |
48.97 |
|
hsa-miR-1260b |
2060.3 |
540.9 |
1270.6 |
1977.9 |
0.26 |
0.43 |
0.27 |
|
hsa-miR-4286 |
678.8 |
202.4 |
2019.2 |
471.2 |
0.30 |
0.10 |
0.43 |

图1.从HeLa细胞中回收的各microRNA量的比较
从人和小鼠细胞株中回收microRNA
通过定量PCR法(miR-92a量)比较使用本产品和原产品从人K562细胞(1×107 cells)和小鼠P388D1细胞(2×107 cells)中获取的microRNA量。结果表明,在同等条件下,使用本产品可获得更多的microRNA。(图2、图3)

图2.人K562细胞中纯化microRNA回收量的比较 图3.小鼠P388D1细胞中纯化microRNA回收量的比较
从人和大鼠血浆中回收游离Ago结合microRNA
通过定量PCR法(miR-92a量)比较使用本产品和原产品(microRNAIsolation Kit, Human Ago2或Mouse Ago2)从人和大鼠血浆中中获取的microRNA量。结果表明,在同等条件下,使用本产品可获得更多的microRNA。(图4、图5)

图4.人血浆中纯化microRNA回收量的比较 图5.大鼠血浆中纯化microRNA回收量的比较
从各种动物细胞株中回收microRNA
通过定量PCR法(miR-92a量)测量使用MagCapture™ microRNA Isolation Kit, Human Ago2、Mouse Ago2、Ago1-4从2×106 cells的K562细胞(人)、COS-7细胞(猴)、MDCK细胞(狗)、P388D1细胞(小鼠)、SCC-131细胞(大鼠)中获取的免疫沉淀RNA片段中microRNA量和使用ISOGEN® 从各种细胞中获取的total RNA片段中microRNA量。比较各免疫沉淀RNA片段中所含miR-92a 量和totalRNA片段中所含miR-92a量的相对值可知,本产品可在全部的细胞种类中获取microRNA。(图6)

图6.从各种细胞中获取的microRNA量的比较
◆MagCapture™ microRNA Isolation Kit, Protein G 实验案例
从人和小鼠细胞株中回收microRNA(与原产品的比较)
通过定量PCR法(miR-92a量)比较在使用本产品(MagCapture™ microRNA Isolation Kit, ProteinG)和抗Ago1抗体(2A7)、抗人Ago2抗体(4G8)、抗人Ago3抗体(1C12)从人K562细胞(1×107 cells)中获得的microRNA量以及使用原产品(microRNA Isolation Kit, Human/Mouse Ago1、Human Ago2、Human Ago3)获得的microRNA量(图1,2,3)。此外,通过定量PCR法(miR-92a量)比较在使用本产品和抗小鼠Ago2抗体(2D4)从小鼠P388D1细胞(2×107 cells)中获得的microRNA量以及使用原产品(microRNA Isolation Kit, Mouse Ago2)获得的microRNA量(图4)。结果表明,本产品和任意抗Ago抗体组合使用,获得的microRNA量要比原产品多。
从人和小鼠细胞株中回收各种Ago蛋白质结合microRNA
通过变性聚丙烯酰胺凝胶电泳和定量PCR法(miR-92a量)研究在使用本产品和抗Ago1抗体(2A7)、抗人Ago3抗体(1C12)、抗Ago4抗体(2G7)从人K562細胞(1×107cells)中获得的microRNA量(图5)。此外,通过变性聚丙烯酰胺凝胶电泳和定量PCR法(miR-92a量)研究在使用本产品和抗Ago1抗体(2A7)、抗小鼠Ago2抗体(2D4)、抗小鼠Ago3抗体(S09)、抗Ago4抗体(2G7)从小鼠P388D1细胞中获得的microRNA量(图6)。结果表明,本产品和任意抗Ago抗体组合使用,可获得各种Ago蛋白结合microRNA。
|
|
|
|
图1. 抗Ago3抗体固定时microRNA回收量的比较 本产品:使用抗Ago1抗体(2A7) 原产品:microRNA Isolation Kit, Human、Mouse Ago1 (291-70201) |
图2. 人Ago2抗体固定时microRNA回收量的比较 本产品:使用抗人Ago2抗体(4G8) 原产品:microRNA Isolation Kit, Human Ago2 (292-66701) |
|
|
|
|
图3. 人Ago3抗体固定时microRNA回收量的比较 本产品:使用抗人Ago3抗体(1C12) 原产品:microRNA Isolation Kit, Human Ago3 (297-70301) |
图4. 小鼠Ago2抗体固定时microRNA回收量的比较 本产品:使用抗小鼠Ago2抗体(2D4) 原产品:microRNA Isolation Kit, Human、Mouse Ago2 (292-67301) |





图5. 从人K562细胞中纯化的microRNA回收量的比较和银染色检验

图6. 从小鼠P388D1细胞中纯化的microRNA回收量的比较和银染色检验
点击此处查看单页
点击图片查看单页
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 293-74801 | MagCapture™microRNA分离试剂盒,Ago1-4 MagCapture™ microRNA Isolation Kit, Ago1-4 |
10次 | - |
| 299-74401 | MagCapture™microRNA分离试剂盒,蛋白G MagCapture™ microRNA Isolation Kit, Protein G |
20次 | - |
| 295-74001 | MagCapture™microRNA分离试剂盒,人Ago2 MagCapture™ microRNA Isolation Kit, Human Ago2 |
10次 | - |
| 297-74201 | MagCapture™microRNA分离试剂盒,小鼠Ago2 MagCapture™ microRNA Isolation Kit, Mouse Ago2 |
10次 | - |
DNA & siRNA转染试剂 ScreenFect™ A plus
用于干细胞研究的小分子化合物
- 产品特性
- 相关资料
- Q&A
- 参考文献
 用于干细胞研究的小分子化合物
用于干细胞研究的小分子化合物
Stemgent® 提供一系列独特的小分子–Stemolecules™ ,这类小分子可用于多能干细胞的自我更新和分化的主要信号通路的分析和调节,诱导多能干细胞(iPS)的细胞重编程,诱导和维持干细胞原始状态。
Stemolecules™为保证高纯度(≥98%)进行了严格检测,通过NMR和质谱进行结构验证,并且测试了对干细胞的细胞毒性。Stemolecules™ 通常用于提高iPS细胞的产生和维持多能干细胞的培养。
| Brand | Name | Catalog Number | Size |
| Stemolecule™ | A83-01 | 04-0014 | 2 mg |
| 04-0014-10 | 10 mg | ||
| Stemolecule™ | ALK5 Inhibitor (RepSox) | 04-0015 | 1 mg |
| Stemolecule™ | All-Trans Retinoic Acid | 04-0021 | 100 mg |
| Stemolecule™ | BI-D1870 | 04-0051 | 10 mg |
| Stemolecule™ | BIO | 04-0003 | 2 mg |
| Stemolecule™ | BIX01294 | 04-0002 | 5 mg |
| Stemolecule™ | CHIR99021 | 04-0004 | 2 mg |
| 04-0004-10 | 10 mg | ||
| Stemolecule™ | CHIR99021 in Solution | 04-0004-02 | 2 mg (10 mM) |
| Stemolecule™ | Cyclopamine | 04-0022 | 2 mg |
| Stemolecule™ | DAPT | 04-0041 | 5 mg |
| Stemolecule™ | Dorsomorphin | 04-0024 | 2 mg |
| Stemolecule™ | Doxycycline hyclate | 04-0016 | 10 mg |
| Stemolecule™ | ec23 | SRP002 | 5 mg |
| SRP002-2 | 2×5mg | ||
| Stemolecule™ | Forskolin | 04-0025 | 10 mg |
| Stemolecule™ | GDC-0941 | 04-0047 | 5 mg |
| Stemolecule™ | GSK429286A | 04-0049 | 2 mg |
| Stemolecule™ | IDE-2 | 04-0027 | 2 mg |
| Stemolecule™ | KAAD-Cyclopamine | 04-0028 | 100 μg |
| Stemolecule™ | LDN-193189 | 04-0074 | 2 mg |
| 04-0074-10 | 10 mg | ||
| Stemolecule™ | LDN-193189 in Solution | 04-0074-02 | 2 mg (10 mM) |
| Stemolecule™ | LY411575 | 04-0054 | 5 mg |
| Stemolecule™ | PD0325901 | 04-0006 | 2 mg |
| 04-0006-10 | 10 mg | ||
| Stemolecule™ | PD0325901 in Solution | 04-0006-02 | 2 mg (10 mM) |
| Stemolecule™ | PD173074 | 04-0008 | 2 mg |
| Stemolecule™ | Purmorphamine | 04-0009 | 5 mg |
| Stemolecule™ | R(+)Bay K 8644 | 04-0013 | 5 mg |
| Stemolecule™ | RG108 | 04-0001 | 10 mg |
| Stemolecule™ | SB431542 | 04-0010 | 5 mg |
| 04-0010-10 | 10 mg | ||
| Stemolecule™ | SB431542 in Solution | 04-0010-05 | 5 mg (10 mM) |
| Stemolecule™ | SB590885 | 04-0080 | 2 mg |
| Stemolecule™ | SC1 (Pluripotin) | 04-0011 | 1 mg |
| Stemolecule™ | SHH Antagonist | 04-0032 | 2 mg |
| Stemolecule™ | Sodium Butyrate | 04-0005 | 500 mg |
| Stemolecule™ | Thiazovivin | 04-0017 | 1 mg |
| Stemolecule™ | Valproic Acid | 04-0007 | 5 g |
| Stemolecule™ | WH-4-023 | 04-0079 | 2 mg |
| Stemolecule™ | Wnt Inhibitor IWP-2 | 04-0034 | 2 mg |
| Stemolecule™ | Wnt Inhibitor IWP-3 | 04-0035 | 2 mg |
| Stemolecule™ | Wnt Inhibitor IWP-4 | 04-0036 | 2 mg |
| Stemolecule™ | XAV939 | 04-0046 | 2 mg |
| Stemolecule™ | Y27632 | 04-0012 | 2 mg |
| 04-0012-10 | 10 mg | ||
| Stemolecule™ | Y27632 Dihydrochloride Hydrate | 04-0012-H-10 | 10 mg |
| Stemolecule™ | Y27632 in Solution | 04-0012-02 | 2 mg (10 mM) |
|
1. |
Chen, S., Liu, Z., Lu, S., & Hu, B. (2019). EPEL promotes the migration and invasion of osteosarcoma cells by upregulating ROCK1. Oncology letters, 17(3), 3133-3140.【StemoleculeTM Y-27632】 |
|
2. |
Zucchini, C., Manara, M. C., Cristalli, C., Carrabotta, M., Greco, S., Pinca, R. S., … & Gambarotti, M. (2019). ROCK2 deprivation leads to the inhibition of tumor growth and metastatic potential in osteosarcoma cells through the modulation of YAP activity. Journal of Experimental & Clinical Cancer Research, 38(1), 1-14. 【StemoleculeTM Y-27632】 |
|
3. |
Cong, F., Hao, H., Hsieh, M. H. L., Jiang, X., Liu, J., & Ng, N. (2019). Cancer patient selection for administration of wnt signaling inhibitors using rnf43 mutation status. U.S. Patent Application No. 16/259,419.【StemoleculeTM Wnt Inhibitor IWP-2】 |
|
4. |
Keller, G., & Craft, A. M. (2019). Keller G, Craft A M. Methods and compositions for generating chondrocyte lineage cells and/or cartilage like tissue. U.S. Patent Application No. 15/977,536. 【StemoleculeTM CHIR99021】 |
|
5. |
Sun, X., Fu, X., Du, M., & Zhu, M. J. (2018). Ex vivo gut culture for studying differentiation and migration of small intestinal epithelial cells. Open biology, 8(4), 170256.【StemoleculeTM DAPT】 |
|
6. |
Gieseck III, R. L., Hannan, N. R., Bort, R., Hanley, N. A., Drake, R. A., Cameron, G. W., … & Vallier, L. (2014). Maturation of induced pluripotent stem cell derived hepatocytes by 3D-culture. PloS one, 9(1), e86372.【Stemolecule CHIR99021】 |
|
7. |
Brick, D. J., Stover, A. E., Nethercott, H. E., Banuelos, M. G., & Schwartz, P. H. (2014). GMP4‐Compatible Production and Expansion of Human Neural Stem Cells. Neural Stem Cell Assays, 35-53. 【StemoleculeTM Y-27632】 |
|
8. |
Fathi, A., Rasouli, H., Yeganeh, M., Salekdeh, G. H., & Baharvand, H. (2015). Efficient differentiation of human embryonic stem cells toward dopaminergic neurons using recombinant LMX1A factor. Molecular biotechnology, 57(2), 184-194. 【StemoleculeTM SB431542,StemoleculeTM LDN-193189】 |
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
ScreenFect™ UP-293 用于重组蛋白大量表达系统的DNA转染试剂
- 产品特性
- 相关资料
- Q&A
- 参考文献
用于重组蛋白大量表达系统的DNA转染试剂
ScreenFect™ UP-293
本产品是适用于FreeStyle293 Expression System和Expi293 Expression System的转染试剂,适用于大量表达重组抗体和蛋白。转染后添加ScreenFect™ UP-293 Booster,可大幅度提高细胞中蛋白的表达量。
◆特点
● 性价比高
● 高重组蛋白表达量
◆试剂盒内容
|
组成试剂 |
100 mL用 |
1 L用 |
|
ScreenFect™ UP-293 Transfection Reagent |
200 μL×1瓶 |
1 mL×2瓶 |
|
ScreenFect™ UP-293 Dilution Buffer |
6.7 mL×1瓶 |
67 mL×1瓶 |
|
ScreenFect™ UP-293 Booster |
500 μL×1瓶 |
5 mL×1瓶 |
本产品仅含转染试剂,不含细胞及培养基。
◆所需其他试剂
|
FreeStyle293 Expression System |
|
|
细胞 |
FreeStyle293-F Cells |
|
培养基 |
FreeStyle293 Expression Medium |
|
稀释缓冲液 |
Opti-MEMI Reduced Serum Medium |
|
Expi293 Expression System |
|
|
细胞 |
Expi293F Cells |
|
培养基 |
Expi293 Expression Medium |
|
稀释缓冲液 |
不需要 |
*用ScreenFect™ UP-293附带的Dilution Buffer稀释。
◆数据
■荧光素酶表达量的比较

用各种转染试剂向FreeStyle293-F细胞和Expi293F细胞中导入分泌型Luc表达载体。
培养规模为30 mL,96小时后收集培养上清,然后用荧光素酶检测法检测发光强度。
FreeStyle293-F细胞:与其他公司的A产品相比,荧光素酶表达量较高。
Expi293F细胞:与其他公司的B产品相比,的荧光素酶表达量相近。
※ 本页面产品仅供研究用,研究以外不可使用。
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 294-80201 | ScreenFect™ UP-293 | 100 mL用 | 基因研究用 |
| 290-80203 | ScreenFect™ UP-293 | 1 L用 | 基因研究用 |
Phos-tag™ SDS-PAGE相关产品
- 产品特性
- 相关资料
- Q&A
- 参考文献
 Phos-tag™ SDS-PAGE相关产品
Phos-tag™ SDS-PAGE相关产品
◆Reagents for Phos-tag™ SDS-PAGE gel preparation
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
30 w/v% Acrylamide/Bis Mixed solution(29:1) |
500 mL |
015-25635 |
Ready-to-use"Solution A".30%T, 3.3%C |
|
Agarose H(High-strength type) |
1 g |
315-01203 |
High-strength Agarose has high strength even in a low-agarose environment and is suitable for electrophoretic migration of large nucleic acid fragments. It can be used in a concentration range of 0.2 – 1% and a separation range of 1 – 200 kbp. |
|
10 g |
319-01201 |
||
|
25 g |
317-01202 |
||
|
10% SDS Solution |
100 mL |
311-90271 |
Ready-to-Use "Solution D" |
|
500 mL |
313-90275 |
||
|
Manganese(Ⅱ) Chloride Tetrahydrate, |
25 g |
134-15302 |
for Molecular Biology Please use for preparation of "Solution F" |
|
100 g |
136-15301 |
||
|
Zinc Chloride |
25 g |
268-01902 |
for Molecular Biology. Please use for preparation of "Solution M" |
|
Bis-Tris |
100 g |
345-04741 |
Please use for preparation of "Solution N" |
|
Sodium Hydrogensulfite〈JIS Special Grade〉 |
25 g |
196-01372 |
Please use for preparation of "Solution O" |
|
100 g |
198-01371 |
||
|
Sodium Hydrogensulfite |
100 g |
190-16461 |
|
|
MOPS |
100 g |
345-01804 |
Please use for preparation of "Solution P" |
|
250 g |
341-01801 |
||
|
MOPS〈For Molecular Biology〉 |
100 g |
341-08241 |
|
|
500 g |
343-08245 |
||
|
Separating Gel Buffer Solution (x4) |
250 mL |
192-11041 |
Ready-to-Use mixed solution of“Sol. B”and“Sol. D”for preparation of Resolving Gel. Contains SDS. |
|
Stacking Gel Buffer Solution (x4) |
250 mL |
199-11051 |
Ready-to-Use mixed solution of“Sol.C”and“Sol. D”for preparation of Stacking Gel. Contains SDS. |
|
N,N,N',N'-Tetramethylethylenediamine (TEMED) |
25 mL |
205-06313 |
for Electrophoresis |
|
10 w/v% Ammonium Peroxodisulfate Solution |
25 mL |
019-15922 |
Ready-to-Use“Solution G” |
◆Premixed Buffers
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
Running Buffer Solution (x10) |
1 L |
184-01291 |
Ready-to-Use concentrated "Solution H" |
|
SDS-PAGE 10x Running Buffer |
1 L |
312-90321 |
Ready-to-Use concentrated "Solution H" |
|
5 L |
318-90323 |
||
|
SDS-PAGE Buffer, pH 8.5 |
5 L |
192-16801 |
Ready-to-Use "Solution H", 1×buffer. |
|
Tricine Running Buffer Solution (×10) |
1 L |
200-17071 |
Composition: 0.5 M Tris/0.5 M Tricine/1% SDS |
|
Sample Buffer Solution (2ME+) (×2) |
25 mL |
196-11022 |
Sample buffer for Laemmli SDS-PAGE containing 2-mercaptoethanol |
|
Sample Buffer Solution |
25 mL |
196-16142 |
Laemmli Sample Buffer containing 3-mercapto-1,2-propanediol (non-hazardous chemical) as substitute for 2-ME |
|
AquaBlot™ 10×High Efficiency Transfer Buffer |
1 L |
015-26213 |
Can use it to transfer to a menbrane with high efficience.
|
◆Enzyme for Dephosphorylation
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
Alkaline Phosphatase(for Biochemistry) |
50 U |
018-10693 |
Applicable to dephosphorylation of proteins |
|
100 U |
012-10691 |
◆Reagents for Staining
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
Quick CBB Plus |
250 mL |
174-00553 |
Ready-to-Use Sol. K. Fixing and destaining procedure are not required. |
|
1 L |
178-00551 |
||
|
Quick-CBB |
2 L |
299-50101 |
By mixing staing solution A and B, ready-to-Use Sol. K |
|
Silver Stain MS Kit |
20 tests |
299-58901 |
Modified proteins by glycosilation and/or phosphorylation can be detected sub-nanogram level on the electrophoretic gel. |
|
Silver Stain Kit Wako |
for 10 gels |
299-13841 |
50~100 times more sensitive than CBB method. |
|
Silver Stain Ⅱ Kit Wako |
for 10 gels |
291-50301 |
This kit contains Stopper, which can be adjusted the staining strength. |
|
Negative Gel Stain MS Kit |
20 tests |
293-57701 |
Applicable to mass analysis and Western blot |
◆Protein Size Marker
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
WIDE-VIEW™ Prestained Protein Size Marker Ⅲ |
25 μL |
236-02463 |
A recommendable prestained marker used with Phos-tag™ SDS-PAGE because obtained bands are less distorted. |
|
500 μL |
230-02461 |
||
|
500 μL x3 |
234-02464 |
◆Positive Control(for confirmation of separation capacity of Phos-tag™ SDS-PAGE)
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
α-Casein, from BovineMilk, Dephosphorylated |
1 mg |
038-23221 |
Mixture of phosphylated and dephosphorylated α-casein |
|
10 mg |
034-23223 |
◆Electrophoresis Apparatus ・ Precast Gels
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
EasySeparator |
1 unit |
058-07681 |
An electrophoresis tank for SuperSep precast polyacrylamide gels. |
|
SuperSep Ace, 6%, 13 wells |
10 gels |
195-15171 |
Prior to use of SuperSep Phos-tag™ PAGE, please use these as sample confirmation. |
|
SuperSep Ace, 7.5%, 13 wells |
10 gels |
198-14941 |
|
|
SuperSep Ace, 7.5%, 17 wells |
10 gels |
191-14931 |
|
|
SuperSep Ace, 10%, 13 wells |
10 gels |
195-14951 |
|
|
SuperSep Ace, 10%, 17 wells |
10 gels |
192-14961 |
|
|
SuperSep Ace, 12.5%, 13 wells |
10 gels |
199-14971 |
|
|
SuperSep Ace, 12.5%, 17 wells |
10 gels |
196-14981 |
|
|
SuperSep Ace, 15%, 13 wells |
10 gels |
193-14991 |
|
|
SuperSep Ace, 15%, 17 wells |
10 gels |
190-15001 |
◆Reagents for Western Blotting
|
Product Name |
Pkg. Size |
Wako Cat. No. |
Use Application |
|
ImmunoStar LD* |
200 cm2 |
296-69901 |
Highly sensitive (femto gram level) immunoblotting, utilizing detection by enhanced chemiluminescence using a unique luminol derivative L-012 as a substrate. |
|
1,000 cm2 |
292-69903 |
||
|
2,000 cm2 |
290-69904 |
||
|
ImmunoStar Zeta* |
200 cm2 |
291-72401 |
Use for detection of proteins between the middle and low femto gram levels. Has stable, long-lasting luminescence signal. |
|
1,000 cm2 |
297-72403 |
||
|
2,000 cm2 |
295-72404 |
||
|
ImmunoStar Basic* |
200 cm2 |
295-75101 |
A cost-effective, stable and long-lasting product. |
|
2,000 cm2 |
291-75103 |
||
|
5,000 cm2 |
299-75104 |
||
|
Immuno Enhancer |
2 assays |
294-68601 |
Ready-to-Use Immunoreaction Enhancer for Western blotting and ELISA |
|
10 assays |
290-68603 |
||
|
40 assays |
298-68604 |
* : Not available for sales in the US and Europe.
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
Phos-tag™ 琼脂糖枪头 Phos-tag™ Tip
- 产品特性
- 相关资料
- Q&A
- 参考文献
Phos-tag™ 琼脂糖枪头
Phos-tag™ Tip
可特异性捕捉磷酸基团的功能性分子“Phos-tag ™”的磷酸化肽纯化用枪头。
枪头里含有 Phos-tag ™ 琼脂糖,是即开即用的前处理工具,适用于生理状态下低分子量磷酸化分子(核酸和多肽等)的分离和富集。
◆原理
把 Phos-tag™ Tip 装在注射器(购买产品附送)上使用。

Phos-tag ™ Tip 的结构

◆优点、特色
|
● 操作时间少于30分钟 ● 高回收率可重复利用 ● 无需昂贵仪器 ● 缓冲液为生理 pH 条件下 |
◆案例、应用
分离磷酸化肽使用例

Phos-tag™ 系列
磷酸化蛋白新方法!
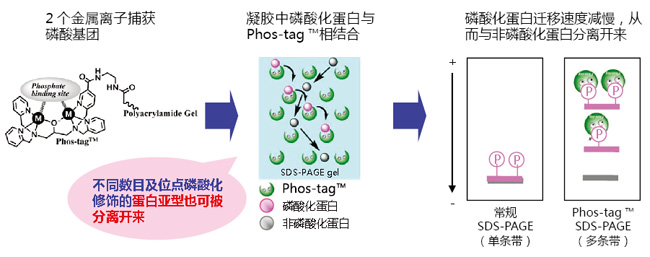
Phos-tag™是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构

◆原理:
◆特点:
● 与 -2 价磷酸根离子的亲和性和选择性高于其它阴离子
● 在 pH 5-8 的生理环境下生成稳定的复合物
◆相关应用:

◆相关产品:
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离:SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离:预制胶中含有 50 μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测:代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化:通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析:用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™ 由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:

Phos-tag 第6版说明书

Phos-tag系列 ver. 8
【参考文献】
· Conversion of graded phosphorylation into switch-like nuclear translocation via autoregulatory mechanisms in ERK signalling[J].Nature communications, 2016, 7,Shindo Y, Iwamoto K, Mouri K, et al.
· PTEN modulates EGFR late endocytic trafficking and degradation by dephosphorylating Rab7[J]. Nature communications, 2016, 7,Shinde S R, Maddika S.
· Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain[J]. Scientific Reports, 2016, 6: 31502,Kawasaki Y, Sakimura A, Park C M, et al.
· Plastid-nucleus communication involves calcium-modulated MAPK signalling[J]. Nature Communications, 2016, 7,Guo H, Feng P, Chi W, et al.
· Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation[J]. Nature communications, 2016, 7,Mitterer V, Murat G, Réty S, et al.
· Phos-tag analysis of Rab10 phosphorylation by LRRK2: a powerful assay for assessing kinase function and inhibitors[J]. Biochemical Journal, 2016: BCJ20160557,Ito G, Katsemonova K, Tonelli F, et al.
· Analysis of phosphorylation of the myosin targeting subunit of smooth muscle myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. The FASEB Journal, 2016, 30(1 Supplement): 1209.1-1209.1,Walsh M P, MacDonald J A, Sutherland C.
· Using Phos-Tag in Western Blotting Analysis to Evaluate Protein Phosphorylation[J]. Kidney Research: Experimental Protocols, 2016: 267-277,Horinouchi T, Terada K, Higashi T, et al.
· The Abundance of Nonphosphorylated Tau in Mouse and Human Tauopathy Brains Revealed by the Use of Phos-Tag Method[J]. The American journal of pathology, 2016, 186(2): 398-409,Kimura T, Hatsuta H, Masuda-Suzukake M, et al.
· Phos-tag SDS-PAGE resolves agonist-and isoform-specific activation patterns for PKD2 and PKD3 in cardiomyocytes and cardiac fibroblasts[J]. Journal of Molecular and Cellular Cardiology, 2016,Qiu W, Steinberg S F.
· Analysis of phosphorylation of the myosin-targeting subunit of myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. American Journal of Physiology-Cell Physiology, 2016, 310(8): C681-C691,Sutherland C, MacDonald J A, Walsh M P.
· Electrochemical biosensor for protein kinase A activity assay based on gold nanoparticles-carbon nanospheres, phos-tag-biotin and β-galactosidase[J]. Biosensors and Bioelectronics, 2016, 86: 508-515,Zhou Y, Yin H, Li X, et al.
· Validation of Cis and Trans Modes in Multistep Phosphotransfer Signaling of Bacterial Tripartite Sensor Kinases by Using Phos-Tag SDS-PAGE[J]. PloS one, 2016, 11(2): e0148294,Kinoshita-Kikuta E, Kinoshita E, Eguchi Y, et al.
· Phosphopeptide Detection with Biotin-Labeled Phos-tag[J]. Phospho-Proteomics: Methods and Protocols, 2016: 17-29,Kinoshita-Kikuta E, Kinoshita E, Koike T.
· A Phos‐tag SDS‐PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1[J]. Proteomics, 2016,Kinoshita E, Kinoshita‐Kikuta E, Kubota Y, et al.
· Difference gel electrophoresis of phosphoproteome: U.S. Patent Application 15/004,339[P]. 2016-1-22,Tao W A, Wang L.
· ERK1/2-induced phosphorylation of R-Ras GTPases stimulates their oncogenic potential[J]. Oncogene, 2016,Frémin C, Guégan J P, Plutoni C, et al.
· Microtubules Inhibit E-Cadherin Adhesive Activity by Maintaining Phosphorylated p120-Catenin in a Colon Carcinoma Cell Model[J]. PloS one, 2016, 11(2): e0148574,Maiden S L, Petrova Y I, Gumbiner B M.
· Serine 231 and 257 of Agamous-like 15 are phosphorylated in floral receptacles[J]. Plant Signaling & Behavior, 2016, 11(7): e1199314,Patharkar O R, Macken T A, Walker J C.
· A small molecule pyrazolo [3, 4-d] pyrimidinone inhibitor of zipper-interacting protein kinase suppresses calcium sensitization of vascular smooth muscle[J]. Molecular pharmacology, 2016, 89(1): 105-117,MacDonald J A, Sutherland C, Carlson D A, et al.
· The RNA polymerase II C-terminal domain phosphatase-like protein FIERY2/CPL1 interacts with eIF4AIII and is essential for nonsense-mediated mRNA decay in Arabidopsis[J]. The Plant Cell, 2016: TPC2015-00771-RA,Chen T, Qin T, Ding F, et al.
· Vasorelaxant Effect of 5′-Methylthioadenosine Obtained from Candida utilis Yeast Extract through the Suppression of Intracellular Ca2+ Concentration in Isolated Rat Aorta[J]. Journal of agricultural and food chemistry, 2016, 64(17): 3362-3370,Kumrungsee T, Akiyama S, Saiki T, et al.
· Inhibition of deubiquitinating activity of USP14 decreases tyrosine hydroxylase phosphorylated at Ser19 in PC12D cells[J]. Biochemical and biophysical research communications, 2016, 472(4): 598-602,Nakashima A, Ohnuma S, Kodani Y, et al.
· Actin Tyrosine-53-Phosphorylation in Neuronal Maturation and Synaptic Plasticity[J]. The Journal of Neuroscience, 2016, 36(19): 5299-5313,Bertling E, Englund J, Minkeviciene R, et al.
· AMPK-dependent phosphorylation of lipid droplet protein PLIN2 triggers its degradation by CMA[J]. Autophagy, 2016, 12(2): 432-438,Kaushik S, Cuervo A M.
· Myocardin-related transcription factor a and yes-associated protein exert dual control in G protein-coupled receptor-and RhoA-mediated transcriptional regulation and cell proliferation[J]. Molecular and cellular biology, 2016, 36(1): 39-49,Olivia M Y, Miyamoto S, Brown J H.
· Extensive phosphorylation of AMPA receptors in neurons[J]. Proceedings of the National Academy of Sciences, 2016, 113(33): E4920-E4927,Diering G H, Heo S, Hussain N K, et al.
· The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis[J]. The Plant Cell, 2016, 28(2): 557-567,Yamamoto Y, Negi J, Wang C, et al.
· The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of molecular and cellular cardiology, 2016, 90: 1-10,Kimura T E, Duggirala A, Smith M C, et al.
· Atg13 is essential for autophagy and cardiac development in mice[J]. Molecular and cellular biology, 2016, 36(4): 585-595,Kaizuka T, Mizushima N.
· The ChrSA and HrrSA two-component systems are required for transcriptional regulation of the hemA promoter in Corynebacterium diphtheriae[J]. Journal of Bacteriology, 2016: JB. 00339-16,Burgos J M, Schmitt M P.
· Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and Immunity, 2016, 84(7): 2086-2093,Zhu L, Olsen R J, Horstmann N, et al.
· Receptor for advanced glycation end products (RAGE) knockout reduces fetal dysmorphogenesis in murine diabetic pregnancy[J]. Reproductive Toxicology, 2016, 62: 62-70,Ejdesjö A, Brings S, Fleming T, et al.
· Aurora kinase-induced phosphorylation excludes transcription factor RUNX from the chromatin to facilitate proper mitotic progression[J]. Proceedings of the National Academy of Sciences, 2016, 113(23): 6490-6495,Chuang L S H, Khor J M, Lai S K, et al.
· Quantitative phosphoproteomics of protein kinase SnRK1 regulated protein phosphorylation in Arabidopsis under submergence[J]. Journal of experimental botany, 2016: erw107,Cho H Y, Wen T N, Wang Y T, et al.
· Temporal regulation of lipin activity diverged to account for differences in mitotic programs[J]. Current Biology, 2016, 26(2): 237-243,Makarova M, Gu Y, Chen J S, et al.
· Block of CDK1‐dependent polyadenosine elongation of Cyclin B mRNA in metaphase‐i‐arrested starfish oocytes is released by intracellular pH elevation upon spawning[J]. Molecular reproduction and development, 2016, 83(1): 79-87,Ochi H, Aoto S, Tachibana K, et al.
· Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1[J]. Cell reports, 2016, 15(9): 2050-2062,Rodriguez-Rodriguez J A, Moyano Y, Játiva S, et al.
· PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells[J]. Cancer medicine, 2016, 5(1): 74-87,Hu Z B, Liao X H, Xu Z Y, et al.
· Phosphorylated TDP-43 becomes resistant to cleavage by calpain: A regulatory role for phosphorylation in TDP-43 pathology of ALS/FTLD[J]. Neuroscience research, 2016, 107: 63-69,Yamashita T, Teramoto S, Kwak S.
· The Pch2 AAA+ ATPase promotes phosphorylation of the Hop1 meiotic checkpoint adaptor in response to synaptonemal complex defects[J]. Nucleic acids research, 2016: gkw506,Herruzo E, Ontoso D, González-Arranz S, et al.
· An optimized guanidination method for large‐scale proteomic studies[J]. Proteomics, 2016,Ye J, Zhang Y, Huang L, et al.
· Expression and purification of the kinase domain of PINK1 in Pichia pastoris[J]. Protein Expression and Purification, 2016,Wu D, Qu L, Fu Y, et al.
· BRI2 and BRI3 are functionally distinct phosphoproteins[J]. Cellular signalling, 2016, 28(1): 130-144,Martins F, Rebelo S, Santos M, et al.
· Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics[J]. Proteomics, 2016,Yang W, Jackson B, Zhang H.
· Regulation of Beclin 1 Protein Phosphorylation and Autophagy by Protein Phosphatase 2A (PP2A) and Death-associated Protein Kinase 3 (DAPK3)[J]. Journal of Biological Chemistry, 2016, 291(20): 10858-10866,Fujiwara N, Usui T, Ohama T, et al.
· Regulatory Implications of Structural Changes in Tyr201 of the Oxygen Sensor Protein FixL[J]. Biochemistry, 2016, 55(29): 4027-4035,Yamawaki T, Ishikawa H, Mizuno M, et al.
· Histone demethylase Jmjd3 regulates osteoblast apoptosis through targeting anti-apoptotic protein Bcl-2 and pro-apoptotic protein Bim[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(4): 650-659,Yang D, Okamura H, Teramachi J, et al.
· Analysis of Molecular Species Profiles of Ceramide-1-phosphate and Sphingomyelin Using MALDI-TOF Mass Spectrometry[J]. Lipids, 2016, 51(2): 263-270,Yamashita R, Tabata Y, Iga E, et al.
· Highly sensitive myosin phosphorylation analysis in the renal afferent arteriole[J]. Journal of Smooth Muscle Research, 2016, 52(0): 45-55,Takeya K.
· Functional dissection of the CroRS two-component system required for resistance to cell wall stressors in Enterococcus faecalis[J]. Journal of bacteriology, 2016, 198(8): 1326-1336,Kellogg S L, Kristich C J.
· Regulation of mitogen-activated protein kinase by protein kinase C and mitogen-activated protein kinase phosphatase-1 in vascular smooth muscle[J]. American Journal of Physiology-Cell Physiology, 2016, 310(11): C921-C930,Trappanese D M, Sivilich S, Ets H K, et al.
· ModProt: a database for integrating laboratory and literature data about protein post-translational modifications[J]. Journal of Electrophoresis, 2016, 60(1): 1-4,Kimura Y, Toda T, Hirano H.
· The C-ETS2-TFEB Axis Promotes Neuron Survival under Oxidative Stress by Regulating Lysosome Activity[J]. Oxidative medicine and cellular longevity, 2016,Ma S, Fang Z, Luo W, et al.
· Essential role of the PSI–LHCII supercomplex in photosystem acclimation to light and/or heat conditions by state transitions[J]. Photosynthesis Research, 2016: 1-10,Marutani Y, Yamauchi Y, Higashiyama M, et al.
· Identification of a redox-modulatory interaction between selenoprotein W and 14-3-3 protein[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(1): 10-18,Jeon Y H, Ko K Y, Lee J H, et al.
· Effects of hydrogen sulfide on the heme coordination structure and catalytic activity of the globin-coupled oxygen sensor AfGcHK[J]. BioMetals, 2016, 29(4): 715-729,Fojtikova V, Bartosova M, Man P, et al.
· Identification and functional analysis of phosphorylation in Newcastle disease virus phosphoprotein[J]. Archives of virology, 2016: 1-14,Qiu X, Zhan Y, Meng C, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports, 2016, 6: 54-62,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· Profiling DNA damage-induced phosphorylation in budding yeast reveals diverse signaling networks[J]. Proceedings of the National Academy of Sciences, 2016: 201602827,Zhou C, Elia A E H, Naylor M L, et al.
· Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system[J]. Nucleic Acids Research, 2016: gkw642,Brosse A, Korobeinikova A, Gottesman S, et al.
· Evolution of ZnII–Macrocyclic Polyamines to Biological Probes and Supramolecular Assembly[J]. Macrocyclic and Supramolecular Chemistry: How Izatt-Christensen Award Winners Shaped the Field, 2016: 415,Kimura E, Koike T, Aoki S.
· Phosphopeptide Enrichment Using Various Magnetic Nanocomposites: An Overview[J]. Phospho-Proteomics: Methods and Protocols, 2016: 193-209,Batalha Í L, Roque A C A.
· In vivo phosphorylation of a peptide tag for protein purification[J]. Biotechnology letters, 2016, 38(5): 767-772,Goux M, Fateh A, Defontaine A, et al.
· Regulation of cell reversal frequency in Myxococcus xanthus requires the balanced activity of CheY‐like domains in FrzE and FrzZ[J]. Molecular microbiology, 2016,Kaimer C, Zusman D R.
· Elevation of cortical serotonin transporter activity upon peripheral immune challenge is regulated independently of p38 mitogen‐activated protein kinase activation and transporter phosphorylation[J]. Journal of neurochemistry, 2016, 137(3): 423-435,Schwamborn R, Brown E, Haase J.
· The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression[J]. Molecular cell, 2016, 62(4): 532-545,Ewald J C, Kuehne A, Zamboni N, et al.
· Two Degradation Pathways of the p35 Cdk5 (Cyclin-dependent Kinase) Activation Subunit, Dependent and Independent of Ubiquitination[J]. Journal of Biological Chemistry, 2016, 291(9): 4649-4657,Takasugi T, Minegishi S, Asada A, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports. 2016,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· a high‐affinity LCO‐binding protein of Medicago truncatula, interacts with LYK3, a key symbiotic receptor[J]. FEBS letters, 2016, 590(10): 1477-1487,Fliegmann J, Jauneau A, Pichereaux C, et al. LYR3,
· Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability[J]. Molecular Cell, 2016,Spies J, Waizenegger A, Barton O, et al.
· PhostagTM-gel retardation and in situ thylakoid kinase assay for determination of chloroplast protein phosphorylation targets[J]. Endocytobiosis and Cell Research, 2016, 27(2): 62-70,Dytyuk Y, Flügge F, Czarnecki O, et al.
· Luteinizing Hormone Causes Phosphorylation and Activation of the cGMP Phosphodiesterase PDE5 in Rat Ovarian Follicles, Contributing, Together with PDE1 Activity, to the Resumption of Meiosis[J]. Biology of reproduction, 2016: biolreprod. 115.135897,Egbert J R, Uliasz T F, Shuhaibar L C, et al.
· Newby, AC, & Bond, M.(2016). The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of Molecular and Cellular Cardiology, 2016, 90: 1-10,Kimura-Wozniak T, Duggirala A, Smith M C, et al. G.
· Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels[J]. The FEBS Journal, 2016, 283(15): 2911-2928,Toume M, Tani M.
· Regulation of CsrB/C sRNA decay by EIIAGlc of the phosphoenolpyruvate: carbohydrate phosphotransferase system[J]. Molecular microbiology, 2016, 99(4): 627-639,Leng Y, Vakulskas C A, Zere T R, et al.
· The Late S-Phase Transcription Factor Hcm1 Is Regulated through Phosphorylation by the Cell Wall Integrity Checkpoint[J]. Molecular and cellular biology, 2016: MCB. 00952-15,Negishi T, Veis J, Hollenstein D, et al.
· Validation of chemical compound library screening for transcriptional co‐activator with PDZ‐binding motif inhibitors using GFP‐fused transcriptional co‐activator with PDZ‐binding motif[J]. Cancer science, 2016, 107(6): 791-802,Nagashima S, Maruyama J, Kawano S, et al.
· ULK1/2 Constitute a Bifurcate Node Controlling Glucose Metabolic Fluxes in Addition to Autophagy[J]. Molecular cell, 2016, 62(3): 359-370,Li T Y, Sun Y, Liang Y, et al.
· Spatiotemporal dynamics of Oct4 protein localization during preimplantation development in mice[J]. Reproduction, 2016: REP-16-0277,Fukuda A, Mitani A, Miyashita T, et al.
· The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress[J]. Parasites & Vectors, 2016, 9(1): 1,Pastor-Fernández I, Regidor-Cerrillo J, Álvarez-García G, et al.
· Interaction Analysis of a Two-Component System Using Nanodiscs[J]. PloS one, 2016, 11(2): e0149187,Hörnschemeyer P, Liss V, Heermann R, et al.
· Constitutive Activation of PINK1 Protein Leads to Proteasome-mediated and Non-apoptotic Cell Death Independently of Mitochondrial Autophagy[J]. Journal of Biological Chemistry, 2016, 291(31): 16162-16174,Akabane S, Matsuzaki K, Yamashita S, et al.
· p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261[J]. Molecular and cellular biology, 2016, 36(10): 1540-1554,Beenstock J, Melamed D, Mooshayef N, et al.
· Lysophosphatidylcholine acyltransferase 1 protects against cytotoxicity induced by polyunsaturated fatty acids[J]. The FASEB Journal, 2016, 30(5): 2027-2039,Akagi S, Kono N, Ariyama H, et al.
· Characterization of a herpes simplex virus 1 (HSV-1) chimera in which the Us3 protein kinase gene is replaced with the HSV-2 Us3 gene[J]. Journal of virology, 2016, 90(1): 457-473,Shindo K, Kato A, Koyanagi N, et al.
· Generation of phospho‐ubiquitin variants by orthogonal translation reveals codon skipping[J]. FEBS letters, 2016, 590(10): 1530-1542,George S, Aguirre J D, Spratt D E, et al.
· Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris[J]. PLoS Genet, 2016, 12(3): e1005922,Ma P, Mori T, Zhao C, et al.
· Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis[J]. FEMS Yeast Research, 2016, 16(6): fow060,Pérez J, Arcones I, Gómez A, et al.
· Alteration of Antiviral Signalling by Single Nucleotide Polymorphisms (SNPs) of Mitochondrial Antiviral Signalling Protein (MAVS)[J]. PloS one, 2016, 11(3): e0151173,Xing F, Matsumiya T, Hayakari R, et al.
· Arm-in-arm response regulator dimers promote intermolecular signal transduction[J]. Journal of bacteriology, 2016, 198(8): 1218-1229,Baker A W, Satyshur K A, Morales N M, et al.
· The lsh/ddm1 homolog mus-30 is required for genome stability, but not for dna methylation in neurospora crassa[J]. PLoS Genet, 2016, 12(1): e1005790,Basenko E Y, Kamei M, Ji L, et al.
· Fine tuning chloroplast movements through physical interactions between phototropins[J]. Journal of Experimental Botany, 2016: erw265,Sztatelman O, Łabuz J, Hermanowicz P, et al.
· Characterization of the Neospora caninum NcROP40 and NcROP2Fam-1 rhoptry proteins during the tachyzoite lytic cycle[J]. Parasitology, 2016, 143(01): 97-113,Pastor-Fernandez I, Regidor-Cerrillo J, Jimenez-Ruiz E, et al.
· Transcriptional Profile during Deoxycholate-Induced Sporulation in a Clostridium perfringens Isolate Causing Foodborne Illness[J]. Applied and environmental microbiology, 2016, 82(10): 2929-2942,Yasugi M, Okuzaki D, Kuwana R, et al.
· Timely Closure of the Prospore Membrane Requires SPS1 and SPO77 in Saccharomyces cerevisiae[J]. Genetics, 2016: genetics. 115.183939,Paulissen S M, Slubowski C J, Roesner J M, et al.
· DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach[J]. Nucleic Acids Research, 2016: gkw626,Wu K Z L, Wang G N, Fitzgerald J, et al.
· OVATE Family Protein 8 Positively Mediates Brassinosteroid Signaling through Interacting with the GSK3-like Kinase in Rice[J]. PLoS Genet, 2016, 12(6): e1006118,Yang C, Shen W, He Y, et al.
· Epithelial Sel1L is required for the maintenance of intestinal homeostasis[J]. Molecular biology of the cell, 2016, 27(3): 483-490, Sun S, Lourie R, Cohen S B, et al.
· Effect of Sodium Dodecyl Sulfate Concentration on Supramolecular Gel Electrophoresis[J]. ChemNanoMat, 2016,Tazawa S, Kobayashi K, Yamanaka M.
· Intergenic VNTR Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and immunity, 2016: IAI. 00258-16,Zhu L, Olsen R J, Horstmann N, et al.
· Ajuba Phosphorylation by CDK1 Promotes Cell Proliferation and Tumorigenesis[J]. Journal of Biological Chemistry, 2016: jbc. M116. 722751,Chen X, Stauffer S, Chen Y, et al.
· Editorial: International Plant Proteomics Organization (INPPO) World Congress 2014[J]. Frontiers in Plant Science, 2016, 7,Heazlewood J L, Jorrín-Novo J V, Agrawal G K, et al.
· Phosphoinositide kinase signaling controls ER-PM cross-talk[J]. Molecular biology of the cell, 2016, 27(7): 1170-1180,Omnus D J, Manford A G, Bader J M, et al.
· A multiple covalent crosslinked soft hydrogel for bioseparation[J]. Chemical Communications, 2016, 52(15): 3247-3250,Liu Z, Fan L, Xiao H, et al.
· Advances in crop proteomics: PTMs of proteins under abiotic stress[J]. Proteomics, 2016, 16(5): 847-865,Wu X, Gong F, Cao D, et al.
· Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae[J]. Molecular cell, 2016, 62(4): 546-557,Zhao G, Chen Y, Carey L, et al.
· Carbon Monoxide Gas Is Not Inert, but Global, in Its Consequences for Bacterial Gene Expression, Iron Acquisition, and Antibiotic Resistance[J]. Antioxidants & redox signaling, 2016,Wareham L K, Begg R, Jesse H E, et al.
· Two-layer regulation of PAQR3 on ATG14-linked class III PtdIns3K activation upon glucose starvation[J]. Autophagy, 2016: 1-2,Xu D, Wang Z, Chen Y.
· Regulation of sphingolipid biosynthesis by the morphogenesis checkpoint kinase Swe1[J]. Journal of Biological Chemistry, 2016, 291(5): 2524-2534,Chauhan N, Han G, Somashekarappa N, et al.
· PAX5 tyrosine phosphorylation by SYK co-operatively functions with its serine phosphorylation to cancel the PAX5-dependent repression of BLIMP1: A mechanism for antigen-triggered plasma cell differentiation[J]. Biochemical and biophysical research communications, 2016, 475(2): 176-181,Inagaki Y, Hayakawa F, Hirano D, et al.
· A Combined Computational and Genetic Approach Uncovers Network Interactions of the Cyanobacterial Circadian Clock[J]. Journal of Bacteriology, 2016: JB. 00235-16,Boyd J S, Cheng R R, Paddock M L, et al.
· HuR mediates motility of human bone marrow-derived mesenchymal stem cells triggered by sphingosine 1-phosphate in liver fibrosis[J]. Journal of Molecular Medicine, 2016: 1-14,Chang N, Ge J, Xiu L, et al.
· Combined replacement effects of human modified β-hexosaminidase B and GM2 activator protein on GM2 gangliosidoses fibroblasts[J]. Biochemistry and Biophysics Reports, 2016,Kitakaze K, Tasaki C, Tajima Y, et al.
· Roseotoxin B Improves Allergic Contact Dermatitis through a Unique Anti-inflammatory Mechanism Involving Excessive Activation of Autophagy in Activated T-Lymphocytes[J]. Journal of Investigative Dermatology, 2016,Wang X, Hu C, Wu X, et al.
References on Phos-tag™ Chemistry
-
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule, Rapid Communications of Mass Spectrometry, 17, 2075-2081 (2003), H. Takeda, A. Kawasaki, M. Takahashi, A. Yamada, and T. Koike
-
Recognition of phosphate monoester dianion by an alkoxide-bridged dinuclear zinc (II) complex, Dalton Transactions, 1189-1193 (2004), E. Kinoshita, M. Takahashi, H. Takeda, M. Shiro, and T. Koike
-
Quantitative analysis of lysophosphatidic acid by time-of-flight mass spectrometry using a phosphate capture molecule, Journal of Lipid Research, 45, 2145-2150 (2004), T. Tanaka, H. Tsutsui, K. Hirano, T. Koike, A. Tokumura, and K. Satouchi
-
Production of 1,2-Didocosahexaenoyl Phosphatidylcholine by Bonito Muscle Lysophosphatidylcholine/Transacylase, Journal of Biochemistry,136, 477-483 (2004), K. Hirano, H. Matsui, T. Tanaka, F. Matsuura, K. Satouchi, and T. Koike
-
Novel immobilized zinc(II) affinity chromatography for phosphopeptides and phosphorylated proteins, Journal of Separation Science, 28, 155-162 (2005), E. Kinoshita, A. Yamada, H. Takeda, E. Kinoshita-Kikuta, and T. Koike
-
Detection and Quantification of On-Chip Phosphorylated Peptides by Surface Plasmon Resonance Imaging Techniques Using a Phosphate Capture Molecule, Analytical Chemistry, 77, 3979-3985 (2005), K. Inamori, M. Kyo, Y. Nishiya, Y. Inoue, T. Sonoda, E. Kinoshita, T. Koike, and Y. Katayama
-
Phosphate-binding tag: A new tool to visualize phosphorylated proteins, Molecular & Cellular Proteomics, 5, 749-757 (2006), E. Kinoshita, E. Kinoshita-Kikuta, K. Takiyama, and T. Koike
-
Enrichment of phosphorylated proteins from cell lysate using phosphate-affinity chromatography at physiological pH, Proteomics, 6, 5088-5095 (2006), E. Kinoshita-Kikuta, E. Kinoshita, A. Yamada, M. Endo, and T. Koike
-
Separation of a phosphorylated histidine protein using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 360, 160-162 (2007), S. Yamada, H. Nakamura, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and Y. Shiro
-
Label-free kinase profiling using phosphate-affinity polyacrylamide gel electrophresis, Molecular & Cellular Proteomics, 6, 356-366 (2007), E. Kinoshita-Kikuta, Y. Aoki, E. Kinoshita, and T. Koike
-
A SNP genotyping method using phosphate-affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 361, 294-298 (2007), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike (The phosphate group at DNA-terminal is efficiently captured by Zn2+.Phos-tag.)
-
Identification on Membrane and Characterization of Phosphoproteins Using an Alkoxide-Bridged Dinuclear Metal Complex as a Phosphate-Binding Tag Molecule, Journal of Biomolecular Techniques, 18, 278-286 (2007), T. Nakanishi, E. Ando, M. Furuta, E. Kinoshita, E. Kikuta-Kinoshita, T. Koike, S. Tsunasawa, and O. Nishimura
-
A mobility shift detection method for DNA methylation analysis using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 378, 102-104 (2008), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Separation of phosphoprotein isotypes having the same number of phosphate groups using phosphate- affinity SDS-PAGE, Proteomics, 8, 2994-3003 (2008), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, S. Yamada, H. Nakamura, Y. Shiro, Y. Aoki, K. Okita, and T. Koike
-
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway, Nature Structural & Molecular Biology, 15, 1138-1146 (2008), M. Ishiai, H. Kitao, A. Smogorzewska, J. Tomida, A. Kinomura, E. Uchida, A. Saberi, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, S. Tashiro, S. J. Elledge, and M. Takata
-
Two-dimensional phosphate affinity gel electrophoresis for the analysis of phosphoprotein isotypes , Electrophoresis, 30, 550-559 (2009), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, Y. Aoki, S. Ohie, Y. Mouri, and T. Koike
-
Formation of lysophosphatidic acid, a wound-healing lipid, during digestion of cabbage leaves, Bioscience, Biotechnology, and Biochemistry,73, 1293-300 (2009), T. Tanaka, G. Horiuchi, M. Matsuoka, K. Hirano, A. Tokumura, T. Koike, and K. Satouchi
-
A Phos-tag-based fluorescence resonance energy transfer system for the analysis of the dephosphorylation of phosphopeptides, Analytical Biochemistry, 388, 235-241, (2009), K. Takiyama, E. Kinoshita, E. Kinoshita-Kikuta, Y. Fujioka, Y. Kubo, and T. Koike
-
Phos-tag beads as an immunoblotting enhancer for selective detection of phosphoproteins in cell lysates, Analytical Biochemistry, 389, 83-85, (2009), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Mobility shift detection of phosphorylation on large proteins using a Phos-tag SDS-PAGE gel strengthened with agarose, Proteomics, 9, 4098- 4101 (2009), E. Kinoshita, E. Kinoshita-Kikuta, H. Ujihara, and T. Koike
-
Separation and detection of large phosphoproteins using Phos-tag SDS-PAGE, Nature Protocols, 4, 1513-1521 (2009), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike
-
A clean-up technology for the simultaneous determination of lysophosphatidic acid and sphingosine-1-phosphate by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a phosphate-capture molecule, Phos-tag, Rapid Communications in Mass Spectrometry, 24, 1075-1084 (2010), J. Morishige, M. Urikura, H. Takagi, K. Hirano, T. Koike, T. Tanaka, and K. Satouchi
-
Genotyping and mapping assay of single-nucleotide polymorphisms in CYP3A5 using DNA-binding zinc(II) complexes, Clinical Biochemistry, 43, 302-306 (2010), E. Kinoshita, E. Kinoshita-Kikuta, H. Nakashima, and T. Koike
-
The DNA-binding activity of mouse DNA methyltransferase 1 is ragulated phosphorylation with casein kinase 1σ/ε, Biochemical Journal, 427, 489-497 (2010), Y. Sugiyama, N. Hatano, N. Sueyoshi, I. Suetake, S. Tajima, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and I. Kameshita
to Page top
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 387-07321 | Phos-tag™ Tip Phos-tag™ 琼脂糖枪头 |
8个 | – |
Phos-tag™ 质谱分析试剂盒 Phos-tag™ Mass Analytical Kit
- 产品特性
- 相关资料
- Q&A
- 参考文献
Phos-tag™ Mass Analytical Kit
用于 MALDI-TOF/MS检测,提高检测灵敏度!
用于质谱分析的试剂套装。
Phos-tag Mass Analytical Kit 是用于质谱分析的试剂套装,可配套 MALDI-TOF/MS 使用。可检测磷酸化分子- Phos-tag® 复合物,通常可提高低磷酸化分子的检测灵敏度。
试剂盒内容:
● Phos-tag™ MS-101L 5 mg([C27H29N6O64Zn2]3+ MW:581.4)
● Phos-tag™ MS-101H 5 mg([C27H29N6O68Zn2]3+ MW:589.4)
● Phos-tag™ MS-101N 10 mg([C27H29N6OZn2]3+ MW:584.3)
◆原理:

◆优点、特色:
● CH3COO- 等价结合在 Phos-tag™ MS-101 上。
● 在溶液中,不含有阴离子的 Phos-tag™ MS-101 带有+3价。
● 检测前需制备 1 mM 的 Phos-tag™ MS-101L,MS-101H 或者 MS-101N(溶于水)。
◆案例、应用:
【使用例子:检测 Phos-tag™ – 磷酸化 LPA 复合体】

由于正电荷增大磷酸化 LPA 检测灵敏度上升
Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™ Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构:

特点:
与-2价磷酸根离子的亲和性和选择性高于其它阴离子
在 pH 5-8 的生理环境下生成稳定的复合物
◆原理:

◆相关应用:

◆相关产品:
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离:SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离:预制胶中含有 50 μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测:代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化:通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析:用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™ 由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:http://phos-tag.jp

Phos-tag 第6版说明书

Phos-tag系列 ver. 8
Q. Phos-tag™ Mass 用于实验可以使用多少次?
A. 如果每次用量为 5 μL,至少可以使用 1000 次。
Q. 如何选择使用 Phos-tag™ MS-101L,Phos-tag™ MS-101H 和 Phos-tag™ MS-101N ?
A. Phos-tag™ 101N 含有自然存在的 Zn,101L 与 101H 分别含有 Zn 的同位素 64Zn 和 68Zn。
请参考以下建议:
摸索条件时使用 101N,其中含有多种同位素,结果比较详细;
鉴定磷酸基团是否存在,使用 101L 和 101H,这些试剂分别包含 64Zn 和 68Zn。使用这些试剂检测同一个样品时会产生不同的荷质比。
Q. 如果想测定经过 Phos-tag™ SDS-PAGE 分离得到的样品,是否必须要在凝胶消化之前去除 Phos-tag™?
A. 没有必要。SDS-PAGE 结束之后根据一般的凝胶消化方法进行操作即可。
Q. 能否用于 ESI 质谱?
A. 是的,可以使用。请参考下面的文献,这篇报道使用 Phos-tag™ MS-101N 进行 ESI-MS 分析。
在实验过程中,使用了中性溶液,若为酸性溶液会导致 Phos-tag ™ 分离。
【参考文献】 Anal. Chem. (2008), 80, 2531-2538 (MS-101N ESI-MS)
【参考文献】
· Conversion of graded phosphorylation into switch-like nuclear translocation via autoregulatory mechanisms in ERK signalling[J].Nature communications, 2016, 7,Shindo Y, Iwamoto K, Mouri K, et al.
· PTEN modulates EGFR late endocytic trafficking and degradation by dephosphorylating Rab7[J]. Nature communications, 2016, 7,Shinde S R, Maddika S.
· Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain[J]. Scientific Reports, 2016, 6: 31502,Kawasaki Y, Sakimura A, Park C M, et al.
· Plastid-nucleus communication involves calcium-modulated MAPK signalling[J]. Nature Communications, 2016, 7,Guo H, Feng P, Chi W, et al.
· Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation[J]. Nature communications, 2016, 7,Mitterer V, Murat G, Réty S, et al.
· Phos-tag analysis of Rab10 phosphorylation by LRRK2: a powerful assay for assessing kinase function and inhibitors[J]. Biochemical Journal, 2016: BCJ20160557,Ito G, Katsemonova K, Tonelli F, et al.
· Analysis of phosphorylation of the myosin targeting subunit of smooth muscle myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. The FASEB Journal, 2016, 30(1 Supplement): 1209.1-1209.1,Walsh M P, MacDonald J A, Sutherland C.
· Using Phos-Tag in Western Blotting Analysis to Evaluate Protein Phosphorylation[J]. Kidney Research: Experimental Protocols, 2016: 267-277,Horinouchi T, Terada K, Higashi T, et al.
· The Abundance of Nonphosphorylated Tau in Mouse and Human Tauopathy Brains Revealed by the Use of Phos-Tag Method[J]. The American journal of pathology, 2016, 186(2): 398-409,Kimura T, Hatsuta H, Masuda-Suzukake M, et al.
· Phos-tag SDS-PAGE resolves agonist-and isoform-specific activation patterns for PKD2 and PKD3 in cardiomyocytes and cardiac fibroblasts[J]. Journal of Molecular and Cellular Cardiology, 2016,Qiu W, Steinberg S F.
· Analysis of phosphorylation of the myosin-targeting subunit of myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. American Journal of Physiology-Cell Physiology, 2016, 310(8): C681-C691,Sutherland C, MacDonald J A, Walsh M P.
· Electrochemical biosensor for protein kinase A activity assay based on gold nanoparticles-carbon nanospheres, phos-tag-biotin and β-galactosidase[J]. Biosensors and Bioelectronics, 2016, 86: 508-515,Zhou Y, Yin H, Li X, et al.
· Validation of Cis and Trans Modes in Multistep Phosphotransfer Signaling of Bacterial Tripartite Sensor Kinases by Using Phos-Tag SDS-PAGE[J]. PloS one, 2016, 11(2): e0148294,Kinoshita-Kikuta E, Kinoshita E, Eguchi Y, et al.
· Phosphopeptide Detection with Biotin-Labeled Phos-tag[J]. Phospho-Proteomics: Methods and Protocols, 2016: 17-29,Kinoshita-Kikuta E, Kinoshita E, Koike T.
· A Phos‐tag SDS‐PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1[J]. Proteomics, 2016,Kinoshita E, Kinoshita‐Kikuta E, Kubota Y, et al.
· Difference gel electrophoresis of phosphoproteome: U.S. Patent Application 15/004,339[P]. 2016-1-22,Tao W A, Wang L.
· ERK1/2-induced phosphorylation of R-Ras GTPases stimulates their oncogenic potential[J]. Oncogene, 2016,Frémin C, Guégan J P, Plutoni C, et al.
· Microtubules Inhibit E-Cadherin Adhesive Activity by Maintaining Phosphorylated p120-Catenin in a Colon Carcinoma Cell Model[J]. PloS one, 2016, 11(2): e0148574,Maiden S L, Petrova Y I, Gumbiner B M.
· Serine 231 and 257 of Agamous-like 15 are phosphorylated in floral receptacles[J]. Plant Signaling & Behavior, 2016, 11(7): e1199314,Patharkar O R, Macken T A, Walker J C.
· A small molecule pyrazolo [3, 4-d] pyrimidinone inhibitor of zipper-interacting protein kinase suppresses calcium sensitization of vascular smooth muscle[J]. Molecular pharmacology, 2016, 89(1): 105-117,MacDonald J A, Sutherland C, Carlson D A, et al.
· The RNA polymerase II C-terminal domain phosphatase-like protein FIERY2/CPL1 interacts with eIF4AIII and is essential for nonsense-mediated mRNA decay in Arabidopsis[J]. The Plant Cell, 2016: TPC2015-00771-RA,Chen T, Qin T, Ding F, et al.
· Vasorelaxant Effect of 5′-Methylthioadenosine Obtained from Candida utilis Yeast Extract through the Suppression of Intracellular Ca2+ Concentration in Isolated Rat Aorta[J]. Journal of agricultural and food chemistry, 2016, 64(17): 3362-3370,Kumrungsee T, Akiyama S, Saiki T, et al.
· Inhibition of deubiquitinating activity of USP14 decreases tyrosine hydroxylase phosphorylated at Ser19 in PC12D cells[J]. Biochemical and biophysical research communications, 2016, 472(4): 598-602,Nakashima A, Ohnuma S, Kodani Y, et al.
· Actin Tyrosine-53-Phosphorylation in Neuronal Maturation and Synaptic Plasticity[J]. The Journal of Neuroscience, 2016, 36(19): 5299-5313,Bertling E, Englund J, Minkeviciene R, et al.
· AMPK-dependent phosphorylation of lipid droplet protein PLIN2 triggers its degradation by CMA[J]. Autophagy, 2016, 12(2): 432-438,Kaushik S, Cuervo A M.
· Myocardin-related transcription factor a and yes-associated protein exert dual control in G protein-coupled receptor-and RhoA-mediated transcriptional regulation and cell proliferation[J]. Molecular and cellular biology, 2016, 36(1): 39-49,Olivia M Y, Miyamoto S, Brown J H.
· Extensive phosphorylation of AMPA receptors in neurons[J]. Proceedings of the National Academy of Sciences, 2016, 113(33): E4920-E4927,Diering G H, Heo S, Hussain N K, et al.
· The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis[J]. The Plant Cell, 2016, 28(2): 557-567,Yamamoto Y, Negi J, Wang C, et al.
· The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of molecular and cellular cardiology, 2016, 90: 1-10,Kimura T E, Duggirala A, Smith M C, et al.
· Atg13 is essential for autophagy and cardiac development in mice[J]. Molecular and cellular biology, 2016, 36(4): 585-595,Kaizuka T, Mizushima N.
· The ChrSA and HrrSA two-component systems are required for transcriptional regulation of the hemA promoter in Corynebacterium diphtheriae[J]. Journal of Bacteriology, 2016: JB. 00339-16,Burgos J M, Schmitt M P.
· Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and Immunity, 2016, 84(7): 2086-2093,Zhu L, Olsen R J, Horstmann N, et al.
· Receptor for advanced glycation end products (RAGE) knockout reduces fetal dysmorphogenesis in murine diabetic pregnancy[J]. Reproductive Toxicology, 2016, 62: 62-70,Ejdesjö A, Brings S, Fleming T, et al.
· Aurora kinase-induced phosphorylation excludes transcription factor RUNX from the chromatin to facilitate proper mitotic progression[J]. Proceedings of the National Academy of Sciences, 2016, 113(23): 6490-6495,Chuang L S H, Khor J M, Lai S K, et al.
· Quantitative phosphoproteomics of protein kinase SnRK1 regulated protein phosphorylation in Arabidopsis under submergence[J]. Journal of experimental botany, 2016: erw107,Cho H Y, Wen T N, Wang Y T, et al.
· Temporal regulation of lipin activity diverged to account for differences in mitotic programs[J]. Current Biology, 2016, 26(2): 237-243,Makarova M, Gu Y, Chen J S, et al.
· Block of CDK1‐dependent polyadenosine elongation of Cyclin B mRNA in metaphase‐i‐arrested starfish oocytes is released by intracellular pH elevation upon spawning[J]. Molecular reproduction and development, 2016, 83(1): 79-87,Ochi H, Aoto S, Tachibana K, et al.
· Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1[J]. Cell reports, 2016, 15(9): 2050-2062,Rodriguez-Rodriguez J A, Moyano Y, Játiva S, et al.
· PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells[J]. Cancer medicine, 2016, 5(1): 74-87,Hu Z B, Liao X H, Xu Z Y, et al.
· Phosphorylated TDP-43 becomes resistant to cleavage by calpain: A regulatory role for phosphorylation in TDP-43 pathology of ALS/FTLD[J]. Neuroscience research, 2016, 107: 63-69,Yamashita T, Teramoto S, Kwak S.
· The Pch2 AAA+ ATPase promotes phosphorylation of the Hop1 meiotic checkpoint adaptor in response to synaptonemal complex defects[J]. Nucleic acids research, 2016: gkw506,Herruzo E, Ontoso D, González-Arranz S, et al.
· An optimized guanidination method for large‐scale proteomic studies[J]. Proteomics, 2016,Ye J, Zhang Y, Huang L, et al.
· Expression and purification of the kinase domain of PINK1 in Pichia pastoris[J]. Protein Expression and Purification, 2016,Wu D, Qu L, Fu Y, et al.
· BRI2 and BRI3 are functionally distinct phosphoproteins[J]. Cellular signalling, 2016, 28(1): 130-144,Martins F, Rebelo S, Santos M, et al.
· Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics[J]. Proteomics, 2016,Yang W, Jackson B, Zhang H.
· Regulation of Beclin 1 Protein Phosphorylation and Autophagy by Protein Phosphatase 2A (PP2A) and Death-associated Protein Kinase 3 (DAPK3)[J]. Journal of Biological Chemistry, 2016, 291(20): 10858-10866,Fujiwara N, Usui T, Ohama T, et al.
· Regulatory Implications of Structural Changes in Tyr201 of the Oxygen Sensor Protein FixL[J]. Biochemistry, 2016, 55(29): 4027-4035,Yamawaki T, Ishikawa H, Mizuno M, et al.
· Histone demethylase Jmjd3 regulates osteoblast apoptosis through targeting anti-apoptotic protein Bcl-2 and pro-apoptotic protein Bim[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(4): 650-659,Yang D, Okamura H, Teramachi J, et al.
· Analysis of Molecular Species Profiles of Ceramide-1-phosphate and Sphingomyelin Using MALDI-TOF Mass Spectrometry[J]. Lipids, 2016, 51(2): 263-270,Yamashita R, Tabata Y, Iga E, et al.
· Highly sensitive myosin phosphorylation analysis in the renal afferent arteriole[J]. Journal of Smooth Muscle Research, 2016, 52(0): 45-55,Takeya K.
· Functional dissection of the CroRS two-component system required for resistance to cell wall stressors in Enterococcus faecalis[J]. Journal of bacteriology, 2016, 198(8): 1326-1336,Kellogg S L, Kristich C J.
· Regulation of mitogen-activated protein kinase by protein kinase C and mitogen-activated protein kinase phosphatase-1 in vascular smooth muscle[J]. American Journal of Physiology-Cell Physiology, 2016, 310(11): C921-C930,Trappanese D M, Sivilich S, Ets H K, et al.
· ModProt: a database for integrating laboratory and literature data about protein post-translational modifications[J]. Journal of Electrophoresis, 2016, 60(1): 1-4,Kimura Y, Toda T, Hirano H.
· The C-ETS2-TFEB Axis Promotes Neuron Survival under Oxidative Stress by Regulating Lysosome Activity[J]. Oxidative medicine and cellular longevity, 2016,Ma S, Fang Z, Luo W, et al.
· Essential role of the PSI–LHCII supercomplex in photosystem acclimation to light and/or heat conditions by state transitions[J]. Photosynthesis Research, 2016: 1-10,Marutani Y, Yamauchi Y, Higashiyama M, et al.
· Identification of a redox-modulatory interaction between selenoprotein W and 14-3-3 protein[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(1): 10-18,Jeon Y H, Ko K Y, Lee J H, et al.
· Effects of hydrogen sulfide on the heme coordination structure and catalytic activity of the globin-coupled oxygen sensor AfGcHK[J]. BioMetals, 2016, 29(4): 715-729,Fojtikova V, Bartosova M, Man P, et al.
· Identification and functional analysis of phosphorylation in Newcastle disease virus phosphoprotein[J]. Archives of virology, 2016: 1-14,Qiu X, Zhan Y, Meng C, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports, 2016, 6: 54-62,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· Profiling DNA damage-induced phosphorylation in budding yeast reveals diverse signaling networks[J]. Proceedings of the National Academy of Sciences, 2016: 201602827,Zhou C, Elia A E H, Naylor M L, et al.
· Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system[J]. Nucleic Acids Research, 2016: gkw642,Brosse A, Korobeinikova A, Gottesman S, et al.
· Evolution of ZnII–Macrocyclic Polyamines to Biological Probes and Supramolecular Assembly[J]. Macrocyclic and Supramolecular Chemistry: How Izatt-Christensen Award Winners Shaped the Field, 2016: 415,Kimura E, Koike T, Aoki S.
· Phosphopeptide Enrichment Using Various Magnetic Nanocomposites: An Overview[J]. Phospho-Proteomics: Methods and Protocols, 2016: 193-209,Batalha Í L, Roque A C A.
· In vivo phosphorylation of a peptide tag for protein purification[J]. Biotechnology letters, 2016, 38(5): 767-772,Goux M, Fateh A, Defontaine A, et al.
· Regulation of cell reversal frequency in Myxococcus xanthus requires the balanced activity of CheY‐like domains in FrzE and FrzZ[J]. Molecular microbiology, 2016,Kaimer C, Zusman D R.
· Elevation of cortical serotonin transporter activity upon peripheral immune challenge is regulated independently of p38 mitogen‐activated protein kinase activation and transporter phosphorylation[J]. Journal of neurochemistry, 2016, 137(3): 423-435,Schwamborn R, Brown E, Haase J.
· The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression[J]. Molecular cell, 2016, 62(4): 532-545,Ewald J C, Kuehne A, Zamboni N, et al.
· Two Degradation Pathways of the p35 Cdk5 (Cyclin-dependent Kinase) Activation Subunit, Dependent and Independent of Ubiquitination[J]. Journal of Biological Chemistry, 2016, 291(9): 4649-4657,Takasugi T, Minegishi S, Asada A, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports. 2016,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· a high‐affinity LCO‐binding protein of Medicago truncatula, interacts with LYK3, a key symbiotic receptor[J]. FEBS letters, 2016, 590(10): 1477-1487,Fliegmann J, Jauneau A, Pichereaux C, et al. LYR3,
· Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability[J]. Molecular Cell, 2016,Spies J, Waizenegger A, Barton O, et al.
· PhostagTM-gel retardation and in situ thylakoid kinase assay for determination of chloroplast protein phosphorylation targets[J]. Endocytobiosis and Cell Research, 2016, 27(2): 62-70,Dytyuk Y, Flügge F, Czarnecki O, et al.
· Luteinizing Hormone Causes Phosphorylation and Activation of the cGMP Phosphodiesterase PDE5 in Rat Ovarian Follicles, Contributing, Together with PDE1 Activity, to the Resumption of Meiosis[J]. Biology of reproduction, 2016: biolreprod. 115.135897,Egbert J R, Uliasz T F, Shuhaibar L C, et al.
· Newby, AC, & Bond, M.(2016). The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of Molecular and Cellular Cardiology, 2016, 90: 1-10,Kimura-Wozniak T, Duggirala A, Smith M C, et al. G.
· Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels[J]. The FEBS Journal, 2016, 283(15): 2911-2928,Toume M, Tani M.
· Regulation of CsrB/C sRNA decay by EIIAGlc of the phosphoenolpyruvate: carbohydrate phosphotransferase system[J]. Molecular microbiology, 2016, 99(4): 627-639,Leng Y, Vakulskas C A, Zere T R, et al.
· The Late S-Phase Transcription Factor Hcm1 Is Regulated through Phosphorylation by the Cell Wall Integrity Checkpoint[J]. Molecular and cellular biology, 2016: MCB. 00952-15,Negishi T, Veis J, Hollenstein D, et al.
· Validation of chemical compound library screening for transcriptional co‐activator with PDZ‐binding motif inhibitors using GFP‐fused transcriptional co‐activator with PDZ‐binding motif[J]. Cancer science, 2016, 107(6): 791-802,Nagashima S, Maruyama J, Kawano S, et al.
· ULK1/2 Constitute a Bifurcate Node Controlling Glucose Metabolic Fluxes in Addition to Autophagy[J]. Molecular cell, 2016, 62(3): 359-370,Li T Y, Sun Y, Liang Y, et al.
· Spatiotemporal dynamics of Oct4 protein localization during preimplantation development in mice[J]. Reproduction, 2016: REP-16-0277,Fukuda A, Mitani A, Miyashita T, et al.
· The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress[J]. Parasites & Vectors, 2016, 9(1): 1,Pastor-Fernández I, Regidor-Cerrillo J, Álvarez-García G, et al.
· Interaction Analysis of a Two-Component System Using Nanodiscs[J]. PloS one, 2016, 11(2): e0149187,Hörnschemeyer P, Liss V, Heermann R, et al.
· Constitutive Activation of PINK1 Protein Leads to Proteasome-mediated and Non-apoptotic Cell Death Independently of Mitochondrial Autophagy[J]. Journal of Biological Chemistry, 2016, 291(31): 16162-16174,Akabane S, Matsuzaki K, Yamashita S, et al.
· p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261[J]. Molecular and cellular biology, 2016, 36(10): 1540-1554,Beenstock J, Melamed D, Mooshayef N, et al.
· Lysophosphatidylcholine acyltransferase 1 protects against cytotoxicity induced by polyunsaturated fatty acids[J]. The FASEB Journal, 2016, 30(5): 2027-2039,Akagi S, Kono N, Ariyama H, et al.
· Characterization of a herpes simplex virus 1 (HSV-1) chimera in which the Us3 protein kinase gene is replaced with the HSV-2 Us3 gene[J]. Journal of virology, 2016, 90(1): 457-473,Shindo K, Kato A, Koyanagi N, et al.
· Generation of phospho‐ubiquitin variants by orthogonal translation reveals codon skipping[J]. FEBS letters, 2016, 590(10): 1530-1542,George S, Aguirre J D, Spratt D E, et al.
· Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris[J]. PLoS Genet, 2016, 12(3): e1005922,Ma P, Mori T, Zhao C, et al.
· Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis[J]. FEMS Yeast Research, 2016, 16(6): fow060,Pérez J, Arcones I, Gómez A, et al.
· Alteration of Antiviral Signalling by Single Nucleotide Polymorphisms (SNPs) of Mitochondrial Antiviral Signalling Protein (MAVS)[J]. PloS one, 2016, 11(3): e0151173,Xing F, Matsumiya T, Hayakari R, et al.
· Arm-in-arm response regulator dimers promote intermolecular signal transduction[J]. Journal of bacteriology, 2016, 198(8): 1218-1229,Baker A W, Satyshur K A, Morales N M, et al.
· The lsh/ddm1 homolog mus-30 is required for genome stability, but not for dna methylation in neurospora crassa[J]. PLoS Genet, 2016, 12(1): e1005790,Basenko E Y, Kamei M, Ji L, et al.
· Fine tuning chloroplast movements through physical interactions between phototropins[J]. Journal of Experimental Botany, 2016: erw265,Sztatelman O, Łabuz J, Hermanowicz P, et al.
· Characterization of the Neospora caninum NcROP40 and NcROP2Fam-1 rhoptry proteins during the tachyzoite lytic cycle[J]. Parasitology, 2016, 143(01): 97-113,Pastor-Fernandez I, Regidor-Cerrillo J, Jimenez-Ruiz E, et al.
· Transcriptional Profile during Deoxycholate-Induced Sporulation in a Clostridium perfringens Isolate Causing Foodborne Illness[J]. Applied and environmental microbiology, 2016, 82(10): 2929-2942,Yasugi M, Okuzaki D, Kuwana R, et al.
· Timely Closure of the Prospore Membrane Requires SPS1 and SPO77 in Saccharomyces cerevisiae[J]. Genetics, 2016: genetics. 115.183939,Paulissen S M, Slubowski C J, Roesner J M, et al.
· DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach[J]. Nucleic Acids Research, 2016: gkw626,Wu K Z L, Wang G N, Fitzgerald J, et al.
· OVATE Family Protein 8 Positively Mediates Brassinosteroid Signaling through Interacting with the GSK3-like Kinase in Rice[J]. PLoS Genet, 2016, 12(6): e1006118,Yang C, Shen W, He Y, et al.
· Epithelial Sel1L is required for the maintenance of intestinal homeostasis[J]. Molecular biology of the cell, 2016, 27(3): 483-490, Sun S, Lourie R, Cohen S B, et al.
· Effect of Sodium Dodecyl Sulfate Concentration on Supramolecular Gel Electrophoresis[J]. ChemNanoMat, 2016,Tazawa S, Kobayashi K, Yamanaka M.
· Intergenic VNTR Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and immunity, 2016: IAI. 00258-16,Zhu L, Olsen R J, Horstmann N, et al.
· Ajuba Phosphorylation by CDK1 Promotes Cell Proliferation and Tumorigenesis[J]. Journal of Biological Chemistry, 2016: jbc. M116. 722751,Chen X, Stauffer S, Chen Y, et al.
· Editorial: International Plant Proteomics Organization (INPPO) World Congress 2014[J]. Frontiers in Plant Science, 2016, 7,Heazlewood J L, Jorrín-Novo J V, Agrawal G K, et al.
· Phosphoinositide kinase signaling controls ER-PM cross-talk[J]. Molecular biology of the cell, 2016, 27(7): 1170-1180,Omnus D J, Manford A G, Bader J M, et al.
· A multiple covalent crosslinked soft hydrogel for bioseparation[J]. Chemical Communications, 2016, 52(15): 3247-3250,Liu Z, Fan L, Xiao H, et al.
· Advances in crop proteomics: PTMs of proteins under abiotic stress[J]. Proteomics, 2016, 16(5): 847-865,Wu X, Gong F, Cao D, et al.
· Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae[J]. Molecular cell, 2016, 62(4): 546-557,Zhao G, Chen Y, Carey L, et al.
· Carbon Monoxide Gas Is Not Inert, but Global, in Its Consequences for Bacterial Gene Expression, Iron Acquisition, and Antibiotic Resistance[J]. Antioxidants & redox signaling, 2016,Wareham L K, Begg R, Jesse H E, et al.
· Two-layer regulation of PAQR3 on ATG14-linked class III PtdIns3K activation upon glucose starvation[J]. Autophagy, 2016: 1-2,Xu D, Wang Z, Chen Y.
· Regulation of sphingolipid biosynthesis by the morphogenesis checkpoint kinase Swe1[J]. Journal of Biological Chemistry, 2016, 291(5): 2524-2534,Chauhan N, Han G, Somashekarappa N, et al.
· PAX5 tyrosine phosphorylation by SYK co-operatively functions with its serine phosphorylation to cancel the PAX5-dependent repression of BLIMP1: A mechanism for antigen-triggered plasma cell differentiation[J]. Biochemical and biophysical research communications, 2016, 475(2): 176-181,Inagaki Y, Hayakawa F, Hirano D, et al.
· A Combined Computational and Genetic Approach Uncovers Network Interactions of the Cyanobacterial Circadian Clock[J]. Journal of Bacteriology, 2016: JB. 00235-16,Boyd J S, Cheng R R, Paddock M L, et al.
· HuR mediates motility of human bone marrow-derived mesenchymal stem cells triggered by sphingosine 1-phosphate in liver fibrosis[J]. Journal of Molecular Medicine, 2016: 1-14,Chang N, Ge J, Xiu L, et al.
· Combined replacement effects of human modified β-hexosaminidase B and GM2 activator protein on GM2 gangliosidoses fibroblasts[J]. Biochemistry and Biophysics Reports, 2016,Kitakaze K, Tasaki C, Tajima Y, et al.
· Roseotoxin B Improves Allergic Contact Dermatitis through a Unique Anti-inflammatory Mechanism Involving Excessive Activation of Autophagy in Activated T-Lymphocytes[J]. Journal of Investigative Dermatology, 2016,Wang X, Hu C, Wu X, et al.
References on Phos-tag™ Chemistry
-
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule, Rapid Communications of Mass Spectrometry, 17, 2075-2081 (2003), H. Takeda, A. Kawasaki, M. Takahashi, A. Yamada, and T. Koike
-
Recognition of phosphate monoester dianion by an alkoxide-bridged dinuclear zinc (II) complex, Dalton Transactions, 1189-1193 (2004), E. Kinoshita, M. Takahashi, H. Takeda, M. Shiro, and T. Koike
-
Quantitative analysis of lysophosphatidic acid by time-of-flight mass spectrometry using a phosphate capture molecule, Journal of Lipid Research, 45, 2145-2150 (2004), T. Tanaka, H. Tsutsui, K. Hirano, T. Koike, A. Tokumura, and K. Satouchi
-
Production of 1,2-Didocosahexaenoyl Phosphatidylcholine by Bonito Muscle Lysophosphatidylcholine/Transacylase, Journal of Biochemistry,136, 477-483 (2004), K. Hirano, H. Matsui, T. Tanaka, F. Matsuura, K. Satouchi, and T. Koike
-
Novel immobilized zinc(II) affinity chromatography for phosphopeptides and phosphorylated proteins, Journal of Separation Science, 28, 155-162 (2005), E. Kinoshita, A. Yamada, H. Takeda, E. Kinoshita-Kikuta, and T. Koike
-
Detection and Quantification of On-Chip Phosphorylated Peptides by Surface Plasmon Resonance Imaging Techniques Using a Phosphate Capture Molecule, Analytical Chemistry, 77, 3979-3985 (2005), K. Inamori, M. Kyo, Y. Nishiya, Y. Inoue, T. Sonoda, E. Kinoshita, T. Koike, and Y. Katayama
-
Phosphate-binding tag: A new tool to visualize phosphorylated proteins, Molecular & Cellular Proteomics, 5, 749-757 (2006), E. Kinoshita, E. Kinoshita-Kikuta, K. Takiyama, and T. Koike
-
Enrichment of phosphorylated proteins from cell lysate using phosphate-affinity chromatography at physiological pH, Proteomics, 6, 5088-5095 (2006), E. Kinoshita-Kikuta, E. Kinoshita, A. Yamada, M. Endo, and T. Koike
-
Separation of a phosphorylated histidine protein using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 360, 160-162 (2007), S. Yamada, H. Nakamura, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and Y. Shiro
-
Label-free kinase profiling using phosphate-affinity polyacrylamide gel electrophresis, Molecular & Cellular Proteomics, 6, 356-366 (2007), E. Kinoshita-Kikuta, Y. Aoki, E. Kinoshita, and T. Koike
-
A SNP genotyping method using phosphate-affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 361, 294-298 (2007), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike (The phosphate group at DNA-terminal is efficiently captured by Zn2+.Phos-tag.)
-
Identification on Membrane and Characterization of Phosphoproteins Using an Alkoxide-Bridged Dinuclear Metal Complex as a Phosphate-Binding Tag Molecule, Journal of Biomolecular Techniques, 18, 278-286 (2007), T. Nakanishi, E. Ando, M. Furuta, E. Kinoshita, E. Kikuta-Kinoshita, T. Koike, S. Tsunasawa, and O. Nishimura
-
A mobility shift detection method for DNA methylation analysis using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 378, 102-104 (2008), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Separation of phosphoprotein isotypes having the same number of phosphate groups using phosphate- affinity SDS-PAGE, Proteomics, 8, 2994-3003 (2008), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, S. Yamada, H. Nakamura, Y. Shiro, Y. Aoki, K. Okita, and T. Koike
-
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway, Nature Structural & Molecular Biology, 15, 1138-1146 (2008), M. Ishiai, H. Kitao, A. Smogorzewska, J. Tomida, A. Kinomura, E. Uchida, A. Saberi, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, S. Tashiro, S. J. Elledge, and M. Takata
-
Two-dimensional phosphate affinity gel electrophoresis for the analysis of phosphoprotein isotypes , Electrophoresis, 30, 550-559 (2009), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, Y. Aoki, S. Ohie, Y. Mouri, and T. Koike
-
Formation of lysophosphatidic acid, a wound-healing lipid, during digestion of cabbage leaves, Bioscience, Biotechnology, and Biochemistry,73, 1293-300 (2009), T. Tanaka, G. Horiuchi, M. Matsuoka, K. Hirano, A. Tokumura, T. Koike, and K. Satouchi
-
A Phos-tag-based fluorescence resonance energy transfer system for the analysis of the dephosphorylation of phosphopeptides, Analytical Biochemistry, 388, 235-241, (2009), K. Takiyama, E. Kinoshita, E. Kinoshita-Kikuta, Y. Fujioka, Y. Kubo, and T. Koike
-
Phos-tag beads as an immunoblotting enhancer for selective detection of phosphoproteins in cell lysates, Analytical Biochemistry, 389, 83-85, (2009), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Mobility shift detection of phosphorylation on large proteins using a Phos-tag SDS-PAGE gel strengthened with agarose, Proteomics, 9, 4098- 4101 (2009), E. Kinoshita, E. Kinoshita-Kikuta, H. Ujihara, and T. Koike
-
Separation and detection of large phosphoproteins using Phos-tag SDS-PAGE, Nature Protocols, 4, 1513-1521 (2009), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike
-
A clean-up technology for the simultaneous determination of lysophosphatidic acid and sphingosine-1-phosphate by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a phosphate-capture molecule, Phos-tag, Rapid Communications in Mass Spectrometry, 24, 1075-1084 (2010), J. Morishige, M. Urikura, H. Takagi, K. Hirano, T. Koike, T. Tanaka, and K. Satouchi
-
Genotyping and mapping assay of single-nucleotide polymorphisms in CYP3A5 using DNA-binding zinc(II) complexes, Clinical Biochemistry, 43, 302-306 (2010), E. Kinoshita, E. Kinoshita-Kikuta, H. Nakashima, and T. Koike
-
The DNA-binding activity of mouse DNA methyltransferase 1 is ragulated phosphorylation with casein kinase 1σ/ε, Biochemical Journal, 427, 489-497 (2010), Y. Sugiyama, N. Hatano, N. Sueyoshi, I. Suetake, S. Tajima, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and I. Kameshita
to Page top
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 305-93551 | Phos-tag™ Mass Analytical Kit Phos-tag 质谱分析试剂盒 |
1 kit | – |
SuperSep Phos-tag™ 预制胶 SuperSep Phos-tag™
- 产品特性
- 相关资料
- Q&A
- 参考文献
蛋白磷酸化研究的预制胶
SuperSep Phos-tag™
SuperSep Phos-tag™ 是研究蛋白磷酸化的方法,无需特异性磷酸化抗体或者同位素标记。

◆SuperSep Phos-tag™
Phos-tag™ 是一种预制胶,预先加入了 50 μmol/L 的 Phostag™ Acrylamide,打开包装即可直接使用。预制胶中含有锌作为金属离子,在中心凝胶缓冲液中保存稳定性很好,得到的带条结果也很整齐。
磷酸化蛋白和非磷酸化蛋白作为不同条带分离。
分离后,胶可用于考马斯亮蓝染色,免疫印迹和质谱实验。
已公开的验证蛋白列表,请点击
◆Phos-tag™ SDS-PAGE 的原理

◆在 HighWire Search 上搜索到的论文数

◆运用
利用 p35 的丙氨酸突变体确定 Cdk5 激活 p35 的磷酸化位点
p35 常见的磷酸化位点是 Ser8 和 Thr138。但是 Ser8 和 Thr138 位点往往会发生丙氨酸突变,产生3种突变体(Ser8 突变体:S8A,Thr138 突变体:T138A,Ser8 和 Thr138 双突变体:2A)。这3种突变体、野生型 p35、Cdk5 和没有激酶活性的 Cdk5 都来源于 COS-7 细胞。这些细胞裂解液用Phos-tag™ SDS-PAGE 和 Western blotting 进行检测(检测抗体:p35 抗体)。

100 μM Phos-tag ™ 丙烯酰胺, 7.5% 聚丙烯酰胺凝胶
可明确磷酸化位点和条带迁移率的关系!
– 泳道1(条带 L2 和 L4)和泳道5(条带 M1):p35 在 Cdk5 的作用下发生了磷酸化;
– 泳道1(条带 L2 和 L4)和泳道3(条带 L2 和 L4):在无激酶活性 Cdk5 的作用下,大约有一半 p35 蛋白在 Thr138 位点发生磷酸化,同样在 138 位发生突变的 p35 蛋白亦是如此。
– 泳道5 (条带 M1)和泳道6(条带 L3 和 L4):Ser8 和 Thr138 是主要的磷酸化位点;
– 泳道5(条带 M1)、泳道7(条带L1和L2)和泳道8(条带M2):条带 M1 是 Ser8 和T hr138 都发生磷酸化的条带;
条带 M2 是只有 Ser8 磷酸化的条带;条带 L1 和 L2 是只有 Thr138 磷酸化的条带。
※ 条带 L1 和 L3 中的X是不确定哪个位点发生磷酸化的条带;
※ 条带L4是非磷酸化的 p35 。
【参考文献】
▪ Quantitative Measurement of in Vivo Phosphorylation States of Cdk5 Activator p35 by Phos-tag ™ SDS-PAGE. T. Hosokawa, T. Saito, A. Asada, K. Fukunaga, and S. Hisanaga,Mol. Cell. Proteomics, Jun 2010;9: 1133 – 1143.
【结果提供】
理化学研究所 脑科学综合研究中心 回路功能研究核心 记忆功能研究团队 细川智永(Dr. T. Hosokawa)
首都大学东京 理工学研究科 生命科学专业 神经分子功能研究室 久永真市(Dr. S. Hisanaga)
检测含有 Dnmt1 磷酸化激酶的片段

我们可以确定在片段中含有目的激酶!
① 采用亲和色谱法从鼠脑提取液中纯化 GST-Dnmt1(1-290)结合蛋白
② 使用 0.3 M 和 1 M NaCl 的 DNA 纤维素柱洗脱得到目的蛋白
③ GST-Dnmt1(1-290)作为体外激酶实验的反应底物
④ Phos-tag ™ SDS-PAGE 用于Western blotting,确定迁移条带中每个片段的激酶活性
【参考文献】
The DNA-binding activity of mouse DNA methyltransferase 1 is regulated by phosphorylation with casein kinase 1delta/epsilon. Y. Sugiyama, N. Hatano, N. Sueyoshi, I. Suetake, S. Tajima, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and I. Kameshita, Biochem. J., May 2010; 427(3): 489-97.
【结果提供】
高知大学 综合研究中心 生命、功能物质部门 实验实习机器设施 杉山康宪(Dr. Y. Sugiyama)
香川大学 农学部 应用生物科学科 动物功能生化学研究室 龟下勇(Dr. I. Kameshita)
二维电泳中的应用:分析 hnRNP K 磷酸化异构体
小鼠巨噬细胞 J774.1 经 LPS 刺激后,裂解细胞,经过免疫沉淀法分离得到 hnRNP K。在二维电泳中,一维是 IPG 胶,二维是 Phos-tag ™ SDS-PAGE,可分离 hnRNP K 的异构体。利用质谱仪,可以确认不同的点代表不同的亚型或修饰蛋白。

同一个等电点的位置上,不同位点发生磷酸化都可以被区分开来
(例: spots 6 vs. 8 and spots 4 vs. 7)
【参考文献】
▪ Characterization of multiple alternative forms of heterogeneous nuclear ribonucleoprotein K by phosphate-affinity electrophoresis. Y. Kimura, K. Nagata, N Suzuki, R. Yokoyama, Y. Yamanaka, H. Kitamura, H. Hirano, and O. Ohara, Proteomics, Nov 2010; 10(21): 3884-95.
【结果提供】
横滨市立大学 生命纳米系统科学研究科 生物体超分子系统科学专业 木村弥生(Dr. Y. Kimura)、平野久(Dr. H. Hirano)
理化学研究所 RCAI 小原收
◆备注
样品制备:
Phos-tag SDS-PAGE 对于蛋白样品中的杂质非常敏感,尤其是螯合剂,钒酸,无机盐,表面活性剂这类物质。
强烈建议在 Phos-tag SDS-PAGE 之前通过 TCA 沉淀或渗析法降低杂质含量。
转膜前处理:
另一个必须的步骤是在转膜前,用 EDTA 去除凝胶中的金属离子(Mn2+ 或者Zn2+);该步骤可提高蛋白的转膜效率。
● 分别准备 10 mmol/L 含 EDTA 和不含 EDTA 两种 1x transfer buffer。
● 将凝胶浸泡在含 10 mmol/L EDTA 的 1x transfer buffer,至少 20 分钟,温和摇晃。更换新缓冲液,重复3次。
● 将凝胶浸泡在不含 10 mmol/L EDTA 的 1x transfer buffer,10 分钟,温和摇晃。
● 转膜操作*。
* 建议用湿法转膜,以提高转膜效率。

◆质量控制
每一批 SuperSep Phos-tag™,出厂前均根据其产品规格进行测试,以保证可分离磷酸化和非磷酸化蛋白,以及他们的分离成都在正常参数内。
◆保存温度
2-10℃
◆产品信息
用于 Bio-Rad 伯乐电泳仪
|
货号 |
品名 |
电泳仪 |
规格 |
|
198-17981 |
SuperSep™ Phos-tag™ (50μmol/L), 7.5%, 17well, 83×100×3.9mm |
Mini-PROTEAN® Tetra Cell (Bio-Rad Laboratories, Inc.) |
5 块 |
|
195-17991 |
SuperSep™ Phos-tag™ (50μmol/L), 12.5%, 17well, 83×100×3.9mm |
5 块 |
用于 Life Technologies 电泳仪
|
货号 |
品名 |
电泳仪 |
规格 |
|
192-18001 |
SuperSep™ Phos-tag™ (50μmol/L), 7.5%, 17well, 100×100×6.6mm |
XCell SureLock® Mini-Cell (Life Technologies, Inc.) |
5 块 |
|
199-18011 |
SuperSep™ Phos-tag™ (50μmol/L), 12.5%, 17well, 100×100×6.6mm |
5 块 |
用于 Wako EasySeperator 电泳仪
| 产品编号 | 产品 | 凝胶浓度 | 孔数 | 包装 |
| 192-17401 | SuperSep™ Phos-tag™ (50 μmol/L) Phos-tag™ 预制胶50 μmol/L |
6.0% | 13孔 | 5块 |
| 199-17391 | 6.0% | 17孔 | 5块 | |
| 195-17371 | 7.5% | 13孔 | 5块 | |
| 192-17381 | 7.5% | 17孔 | 5块 | |
| 193-16711 | 10.0% | 13孔 | 5块 | |
| 190-16721 | 10.0% | 17孔 | 5块 | |
| 195-16391 | 12.5% | 13孔 | 5块 | |
| 193-16571 | 12.5% | 17孔 | 5块 | |
| 193-16691 | 15.0% | 13孔 | 5块 | |
| 196-16701 | 15.0% | 17孔 | 5块 | |
| 197-16851 | 17.5% | 13孔 | 5块 | |
| 194-16861 | 17.5% | 17孔 | 5块 |
◆相关产品
| 产品编号 | 产品名称 | 规格 |
| 058-07681 | EasySeparator Phos-tag预制凝胶的配套电泳槽 |
1 set |
Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™ Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构:

特点:
与 -2 价磷酸根离子的亲和性和选择性高于其它阴离子
在 pH 5-8 的生理环境下生成稳定的复合物
◆原理:
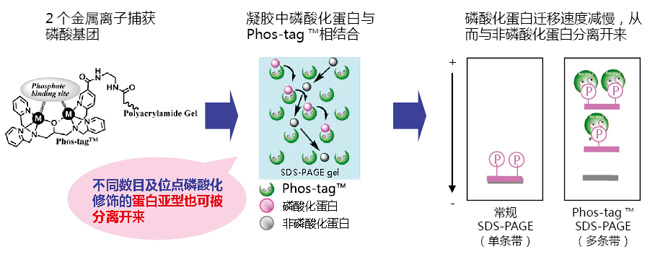
◆相关应用:

◆相关产品:
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离: SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离: 预制胶中含有50μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测: 代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化: 通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析: 用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™ 由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:

Phos-tag 第6版说明书

Phos-tag系列 ver. 8
|
Q1. |
我们可以采用哪种凝胶染色法? |
|
A1. |
所有的染色法都可使用,最常用的例如考马斯亮蓝染色法,负染,银染和荧光染色等。 |
|
Q2. |
用考马斯亮蓝染法染色,着色不明显。 |
|
A2. |
在微波炉内进行脱色,会取得比较满意的效果。 方法:将染色的胶放在 100 毫升去离子水里,用擦拭纸包裹胶,再放进微波炉加热几分钟后更换去离子水,并重复上述步骤3或4次。请注意防止盛放胶的容器温度过高。 |
|
Q3. |
此款产品能否用于免疫印迹? |
|
A3. |
可以,如果用 EDTA 清除胶里面含有的锌,可以提高转膜的效率。 方法:胶浸在含有 10 mmol/L EDTA 的转移缓冲液(25 mmol/L tris、192 mmol/L甘氨酸,10%甲醇)里轻轻搅拌 10 分钟。重复上述步骤3次。然后放进不含 EDTA 的转移缓冲液(25 mmol/L tris、192 mmol/L的甘氨酸,10%甲醇)里搅拌 10 分钟并转移到 PVDF 膜或 NC 膜(硝酸纤维素膜)上。 |
|
Q4. |
条带扭曲了。 |
|
A4. |
含有 EDTA,无机盐,表面活性剂等的样品可能会导致条带弯曲或者拖尾。通过 TCA 或透析沉淀脱盐样品。空白泳道也会导致相邻样品条带弯曲,在空白泳道加与样品相同体积的样品缓冲液(x1)。 |
|
Q5. |
磷酸化蛋白和非磷酸化蛋白不能分离。 |
|
A5. |
将β–酪蛋白(038-23221)作为 Phos-tag SDS-PAGE 电泳的阳性对照,将用碱性磷酸酶处理的β–酪蛋白作为阴性对照,并检查条带的迁移。如果只有一个条带,可能是由于 Phos-tag™ 或丙烯酰胺的浓度没有优化导致磷酸化和非磷酸化蛋白不能分离。 |
|
Q6. |
可用在细胞的粗提物上吗? |
|
A6. |
可以的,可能 Rf 值可能有稍低,由于目的蛋白的因素,条带可能会比较模糊。 |
|
Q7. |
上样量多少? |
|
A7. |
1至5微克纯化的蛋白(考马斯亮蓝染法),10 至 30 微克的组织或细胞提取物(取决于蛋白质的表达量)。 *这是推荐的用量,可以先进行常规的 SDS-PAGE 和免疫印迹法,确定合适的上样量。 |
|
Q8. |
使用哪种蛋白marker? |
|
A8. |
不推荐使用蛋白 marker。该款产品不需要蛋白 marker。因此,建议用来源于大肠杆菌的重组蛋白或者是非磷酸化样品作为阴性对照代替蛋白 marker。 |
|
Q9. |
此款产品的配离子是什么? |
|
A9. |
锌离子 |
|
Q10. |
我们怎么知道是因为蛋白发生磷酸化条带才会发生迁移? |
|
A10. |
使用 12.5% SuperSep™ Ace(Wako 目录 No. 199-14971)进行电泳(有相同的胶浓度),检查目标蛋白质是否降解。 |
参考文献
| 1. |
Apostolidis, S. A., Rauen, T., Hedrich, C. M., Tsokos, G. C., & Crispín, J. C. (2013). Protein phosphatase 2A enables expression of interleukin 17 (IL-17) through chromatin remodeling. Journal of Biological Chemistry, 288(37), 26775-26784. 全文 【IF=4.238】 |
| 2. |
Baek, J. H., Gomez, I. G., Wada, Y., Roach, A., Mahad, D., & Duffield, J. S. (2018). Deletion of the Mitochondrial Complex-IV Cofactor Heme A: Farnesyltransferase Causes Focal Segmental Glomerulosclerosis and Interferon Response. The American journal of pathology, 188(12), 2745-2762. 全文 【IF:3.491,预制胶浓度7.5%】 |
| 3. | Bouvet, M., Dubois-Deruy, E., Alayi, T. D., Mulder, P., El Amranii, M., Beseme, O., … & Pinet, F. (2016). Increased level of phosphorylated desmin and its degradation products in heart failure. Biochemistry and biophysics reports, 6, 54-62. 全文
【预制胶浓度10%】 |
| 4. | Boisvert, R. A., Schuttert, C. W., Shanahan, L. M., & Howlett, N. G. (2015). Characterization of a Putative Cyclin Dependent Kinase Phosphorylation Site Cluster in FANCD2. Further Insights Into the Regulation of the Fanconi Anemia FANCD2 Protein, 106-168. 全文
【预制胶浓度6%】 |
| 5. | Chen, A. H., Lubkowicz, D., Yeong, V., Chang, R. L., & Silver, P. A. (2015). Transplantability of a circadian clock to a noncircadian organism. Science advances, 1(5), e1500358. 全文
【IF:13.116】 |
| 6. |
Chung, S., Kijima, K., Kudo, A., Fujisawa, Y., Harada, Y., Taira, A., … & Nakamura, Y. (2016). Preclinical evaluation of biomarkers associated with antitumor activity of MELK inhibitor. Oncotarget, 7(14), 18171. 全文 【IF:5.168(2016年),预制胶浓度12.5%】 |
| 7. | Ciftci, H. I., Fujino, H., Koga, R., Yamamoto, M., Kawamura, S., Tateishi, H., … & Fujita, M. (2015). Mutational analysis of HIV-2 Vpx shows that proline residue 109 in the poly-proline motif regulates degradation of SAMHD1. FEBS letters, 589(13), 1505-1514. 全文
【IF:3.057,预制胶浓度15%】 |
| 8. | Dupré, E., Lesne, E., Guérin, J., Lensink, M. F., Verger, A., De Ruyck, J., … & Jacob-Dubuisson, F. (2015). Signal transduction by BvgS sensor kinase. Journal of Biological Chemistry, 290(38), 23307-23319. 全文
【IF:4.238,预制胶浓度12.5%】 |
| 9. | Edens, L. J., Dilsaver, M. R., & Levy, D. L. (2017). PKC-mediated phosphorylation of nuclear lamins at a single serine residue regulates interphase nuclear size in Xenopus and mammalian cells. Molecular biology of the cell, 28(10), 1389-1399. 全文
【IF:3.791,预制胶浓度10%】 |
| 10. | Ishibashi, T., Morita, S., Kishimoto, S., Uraki, S., Takeshima, K., Furukawa, Y., … & Nishi, M.(2020). Nicotinic acetylcholine receptor signaling regulates inositol-requiring enzyme 1a activation to protect β-cells against terminal unfolded protein response under irremediable endoplasmic reticulum stress. Journal of Diabetes Investigation, 11(4), 801-813 全文
【IF:3.761】 |
| 11. |
Jakobsen, T. H., Warming, A. N., Vejborg, R. M., Moscoso, J. A., Stegger, M., Lorenzen, F., … & Nielsen, T. E. (2017). A broad range quorum sensing inhibitor working through sRNA inhibition. Scientific reports, 7(1), 1-12. 全文 【IF:3.998,预制胶浓度12.5%】 |
| 12. | Kariya, Y., Kanno, M., Matsumoto-Morita, K., Konno, M., Yamaguchi, Y., & Hashimoto, Y. (2014). Osteopontin O-glycosylation contributes to its phosphorylation and cell-adhesion properties. Biochemical Journal, 463(1), 93-102. 全文
【IF:4.097,预制胶浓度7.5%】 |
| 13. | Kawabata, N., & Matsuda, M. (2016). Cell density-dependent increase in tyrosine-monophosphorylated ERK2 in MDCK cells expressing active Ras or Raf. PloS one, 11(12), e0167940. 全文
【IF:2.74,预制胶浓度5-20%】 |
| 14. |
Kinoshita-Kikuta, E. , Kinoshita, E. , & Koike, T. . (2012). A laborsaving, timesaving, and more reliable strategy for separation of low-molecular-mass phosphoproteins in phos-tag affinity electrophoresis. International Journal of Chemistry, 4(5). 全文 |
| 15. | Kinoshita, E. , Kinoshita-Kikuta, E. , & Koike, T. . (2015). Advances in phos-tag-based methodologies for separation and detection of the phosphoproteome. Biochimica et Biophysica Acta (BBA) – Proteins & Proteomics, 1854(6), 601-608. 全文
【IF:2.371,预制胶浓度12.5%】 |
| 16. | Maria Blaire, Bustamante, Annalisa, Ansaloni, Jeppe Falsig, & Pedersen, et al. (2015). Detection of huntingtin exon 1 phosphorylation by phos-tag SDS-PAGE: predominant phosphorylation on threonine 3 and regulation by ikkβ. Biochemical and biophysical research communications. 全文
【IF:2.985,预制胶浓度12.5%】 |
| 17. |
Najafov, A., Mookhtiar, A. K., Luu, H. S., Ordureau, A., Pan, H., Amin, P. P., … & Yuan, J. (2019). TAM kinases promote necroptosis by regulating oligomerization of MLKL. Molecular cell, 75(3), 457-468. 全文 【IF:15.584】 |
| 18. |
Oku, Y., Nishiya, N., Shito, T., Yamamoto, R., Yamamoto, Y., Oyama, C., & Uehara, Y. (2015). Small molecules inhibiting the nuclear localization of YAP/TAZ for chemotherapeutics and chemosensitizers against breast cancers. FEBS open bio, 5, 542-549. 全文 【IF:2.231】 |
| 19. | Oyama, M., Kariya, Y., Kariya, Y., Matsumoto, K., Kanno, M., Yamaguchi, Y., & Hashimoto, Y. (2018). Biological role of site-specific O-glycosylation in cell adhesion activity and phosphorylation of osteopontin. Biochemical Journal, 475(9), 1583-1595. 全文
【IF:4.097,预制胶浓度7.5%; 5-20%】 |
| 20. |
Pan, L., Hildebrand, K., Stutz, C., Thomä, N., & Baumann, P. (2015). Minishelterins separate telomere length regulation and end protection in fission yeast. Genes & development, 29(11), 1164-1174. 全文 【IF:9.527,预制胶浓度10%】 |
| 21. |
Tsukamoto, S., Kuratani, M., & Katagiri, T. (2020). Functional characterization of a unique mutant of ALK2, p. K400E, that is associated with a skeletal disorder, diffuse idiopathic skeletal hyperostosis. Bone, 115410. 全文 【IF:4.147,预制胶浓度7.5%】 |
| 22. | Sartagul, W., Zhou, X., Yamada, Y., Ma, N., Tanaka, K., Furuyashiki, T., & Ma, Y. (2014). The MluI cell cycle box (MCB) motifs, but not damage-responsive elements (DREs), are responsible for the transcriptional induction of the rhp51+ gene in response to DNA replication stress. PloS one, 9(11), e111936. 全文
【IF:2.74,预制胶浓度10%】 |
| 23. | Tavernier, Q., Bennana, E., Poindessous, V., Schaeffer, C., Rampoldi, L., Pietrancosta, N., & Pallet, N. (2018). Regulation of IRE1 RNase activity by the Ribonuclease inhibitor 1 (RNH1). Cell Cycle, 17(15), 1901-1916. 全文
【IF:3.699】 |
| 24. | Watanabe, A., Sasaki, T., Yukami, T., Kanki, H., Sakaguchi, M., Takemori, H., … & Mochizuki, H. (2016). Serine racemase inhibition induces nitric oxide-mediated neurovascular protection during cerebral ischemia. Neuroscience, 339, 139-149. 全文
【IF:3.056,预制胶浓度12.5%】 |
| 25. | Watanabe, T., Tsuruoka, M., Narita, Y., Katsuya, R., Goshima, F., Kimura, H., & Murata, T. (2015). The Epstein–Barr virus BRRF2 gene product is involved in viral progeny production. Virology, 484, 33-40. 全文
【IF:2.819,预制胶浓度7.5%】 |
| 26. |
Xing, F., Matsumiya, T., Hayakari, R., Yoshida, H., Kawaguchi, S., Takahashi, I., … & Imaizumi, T. (2016). Alteration of antiviral signalling by single nucleotide polymorphisms (SNPs) of mitochondrial antiviral signalling protein (MAVS). PloS one, 11(3), e0151173. 全文 【IF:2.74,预制胶浓度10%】 |
| 27. | Zhang, X., Hu, P., Ding, S. Y., Sun, T., Liu, L., Han, S., … & Wang, X. (2019). Induction of autophagy-dependent apoptosis in cancer cells through activation of ER stress: an uncovered anti-cancer mechanism by anti-alcoholism drug disulfiram. American journal of cancer research, 9(6), 1266-1281. 全文
【IF:5.177,预制胶浓度7.5%】 |
| 28. | Zhao, D., Lu, X., Wang, G., Lan, Z., Liao, W., Li, J., … & Tang, M. (2017). Synthetic essentiality of chromatin remodelling factor CHD1 in PTEN-deficient cancer. Nature, 542(7642), 484-488. 全文
【IF:42.778】 |
| 29. | Funabara, D., Nishimura, Y., & Kanoh, S. (2019). Phosphorylation Properties of the N-Terminal Region of Twitchin from Molluscan Catch Muscle. American Journal of Molecular Biology, 9(3), 110-120. 概要
【预制胶浓度12.5%】 |
| 30. |
都築政弘, 村田真理子, & 再生脳外科学. (2012). 新規染色法○ 董 偉傑・松野裕樹・亀山昭彦. 生物物理化学, 56, s90. 全文 【预制胶浓度12.5%】 |
| 31. | 脇本理恵子, 木下恵美子, 木下英司, & 小池透.(2014). P-1 SuperSep Phos-tagを用いた低分子量リン酸化タパク質の解析. 生物物理化学, Vol 58, 2, 98-108. 全文
【预制胶浓度12.5%】 |
| 32. | Nogami, M., Ishikawa, M., Sano, O., Sone, T., Akiyama, T., Aoki, M., … & Okano, H. Identification of Hub Molecules of FUS-ALA by Bayesian Gene Regulatory Network Analysis of iPSC Model: iBRN. 概要 |
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 193-16711 | SuperSep Phos-tag™ (50 μmol/L), 10%, 13 well Phos-tag 13孔10%预制胶 |
5块 | – |
| 190-16721 | SuperSep Phos-tag™ (50 μmol/L), 10%, 17 well Phos-tag 17孔10%预制胶 |
5块 | – |
| 195-16391 | SuperSep Phos-tag™ (50 μmol/L), 12.5%, 13 well Phos-tag 13孔12.5%预制胶 |
5块 | – |
| 193-16571 | SuperSep Phos-tag™ (50 μmol/L), 12.5%, 17 well Phos-tag 17孔12.5%预制胶 |
5块 | – |
| 193-16691 | SuperSep Phos-tag™ (50 μmol/L), 15%, 13 well Phos-tag 13孔15%预制胶 |
5块 | – |
| 196-16701 | SuperSep Phos-tag™ (50 μmol/L), 15%, 17 well Phos-tag 17孔15%预制胶 |
5块 | – |
| 197-16851 | SuperSep Phos-tag™ (50 μmol/L), 17.5%, 13 well Phos-tag 13孔17.5%预制胶 |
5块 | – |
| 194-16861 | SuperSep Phos-tag™ (50 μmol/L), 17.5%, 17 well Phos-tag 17孔17.5%预制胶 |
5块 | – |
Phos-tag™ 生物素 Phos-tag™ Biotin
- 产品特性
- 相关资料
- Q&A
- 参考文献
Phos-tag™ Biotin
无特异性磷酸化抗体时的理想选择!
Phos-tag™ Biotin 是与生物素结合的 Phos-tag™,可用于免疫印迹法检测磷酸化蛋白。Phos-tag™ Biotin BTL-104 和 BTL-105 可灵敏检测 PVDF 膜上的磷酸化蛋白。
◆原理

◆优点、特色
● 无辐射。
● 无需 PVDF 膜的封闭处理。
● Phos-tag™ 的特异性结合与氨基酸种类、序列无关。
● 可适用于免疫印迹和质谱分析等后续工作。
● Phos-tag™ BTL 母液可稳定保存至少6个月。
● 实验流程与使用 HRP 标记抗体相类似。
※BTL-104、BTL-105、BTL-111 三者连接链(Linker)长度不一,但使用上基本相同。BTL-111 灵敏度更高。
◆案例、应用
【使用例:在 PVDF 膜上检测磷酸化蛋白】

转印在 PVDF 膜上的磷酸化蛋白可精确检测到 ng 级水平,没有检测到相应的去磷酸化蛋白与非磷酸化蛋白的信号斑点。
免疫印迹检测磷酸化蛋白——Phos-tag ™ 生物素。
摘自 Eiji Kinoshita ,et al., Mol.Cel.Proteomics (2006) 5: 749
【使用例:Phos-tag™ 生物素在检测蛋白激酶活性的微阵列(生物芯片)中的运用】
蛋白激酶是很多疾病诊断和药物筛选的靶标。近来有科研人员研发了一种检测胞内蛋白激酶活性的高灵敏度多肽微阵列。用微阵列点样机点样 2 nL 体积的底物多肽溶液,使多肽固定在戊二醛预修饰的高氨基末端载玻片。
当多肽经细胞裂解液磷酸化后,用荧光标记的抗 phosphotyrosine(磷酸化酪氨酸)抗体检测酪氨酸激酶,或者用 Phos-tag™ 生物素,接着用荧光标记的亲和素检测丝氨酸或者苏氨酸激酶。之后用自动微阵列扫描仪检测荧光信号。多肽微阵列系统包括简单的多肽固定,只需少量样品,具有高密度阵列。重要的是,检测细胞裂解液蛋白激酶活性的灵敏度高。
因此多肽微阵列系统可用于高通量筛选细胞内激酶活性,可用于药物筛选和疾病诊断。
Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构

◆特点
与 -2 价磷酸根离子的亲和性和选择性高于其它阴离子
在 pH 5-8 的生理环境下生成稳定的复合物
◆原理
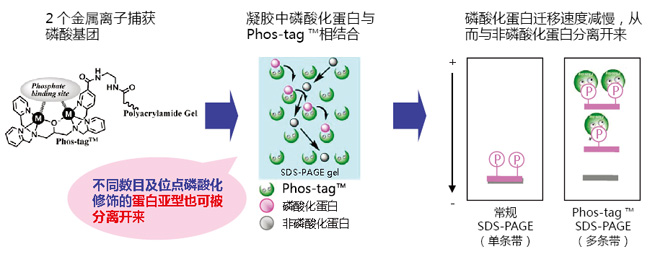
◆相关应用

◆相关产品
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离:SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离:预制胶中含有50 μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测:代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化:通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析:用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:
|
Phos-tag 第6版说明书 |
Phos-tag 生物素操作手册
|
|
说明书 |



Q: BTL-104 和 BTL-111 的区别?
A: BTL-104 的溶解度更高,BTL-111 的灵敏度更高。
Q: 检测灵敏度达到什么水平?
A: 可达到 ng 级别。需要使用化学发光试剂,比如 ImmunoStar LD(Wako)。
Q: 使用该产品还需要别的试剂或则耗材吗?
A: 硝酸锌(Zn(NO3)2)溶液和亲和素标记的 HRP((GE Healthcare Bio-Sciences: RPN1231)。制备 Phos-tag™ 生物素与亲和素标记的 HRP
偶联物时需要使用离心过滤装置(NMWL = 30,000, Nanosep™ 30K, Pall Life Sciences)。
Q: Phos-tag™生物素使用的次数?
A: 主要决定于使用次数和使用量,以下实验次数仅作参考:
BTL-104:130~1300次;
BTL-111 1 mM 溶液:10~100次。
Q: 可测定磷酸化蛋白?
A: 根据条带的浓度,可进行半定量分析。
Q: 能够确定结合磷酸化基团的数目?
A: 不能。
Q: 能否剥除(strip)Phos-tag™生物素?
A: 可以。与含有 62.5 mM Tris-HCl(pH6.8),2%(w/v)SDS 和 0.1M 2-巯基乙醇溶液 混合后,震荡15分钟。用 1×TBS-T 洗涤3次,
每次 10 分钟。
Q: 推荐使用哪种膜?
A: 建议使用 PVDF 膜。
Q: 使用 Phos-tag™ 生物素是否需要封闭?
A: 不需要。封闭会降低检测灵敏度。
【参考文献】
| 1. |
"Phosphate-binding tag: A new tool to visualize phosphorylated proteins", E. Kinoshita, E. Kinoshita-Kikuta, K. Takiyama, and T. Koike, Mol. Cell. Proteomics, 2006, 5, 749-757. |
| 2. |
"Detection and Quantification of On-Chip Phosphorylated Peptides by Surface Plasmon Resonance Imaging Techniques Using a Phosphate Capture Molecule", K. Inamori, M. Kyo, Y. Nishiya, Y. Inoue, T. Sonoda, E. Kinoshita, T. Koike, and Y. Katayama, Anal. Chem., 2006, 77, 3979-3985. |
| 3. |
"Phos-tagを用いたりん酸化タンパク質の検出法 ①ビオチン化Phos-tagを用いたケミルミ検出", 木下英司, 木下恵美子, 小池透, 実験医学, 2006, 24, 2523-2529. |
| 4. |
"Identification on Membrane and Characterization of Phosphoproteins Using an Alkoxide-Bridged Dinuclear Metal Complex as a Phosphate-Binding Tag Molecule", T. Nakanishi, E. Ando, M. Furuta, E. Kinoshita, E. Kikuta-Kinoshita, T. Koike, S. Tsunasawa, and O. Nishimura, J. Biomol. Tech., 2007, 18, 278-286. |
| 5. |
Wang, H. , Qi, C. , He, W. , Wang, M. , Jiang, W. , & Yin, H. , et al. (2018). A sensitive photoelectrochemical immunoassay of n(6)-methyladenosine based on dual-signal amplification strategy: ru doped in sio2 nanosphere and carboxylated g-c3n4. Biosensors & Bioelectronics, 99, 281.【用于m6A检测】 |
| 6. |
Li B , Yin H , Zhou Y , et al. Photoelectrochemical detection of miRNA-319a in rice leaf responding to phytohormones treatment based on CuO-CuWO 4, and rolling circle amplification[J]. Sensors and Actuators B: Chemical, 2017:S0925400517316295. |
| 7. |
Emiko, K. K., Eiji, K., & Tohru, K. (2016). Phosphopeptide detection with biotin-labeled phos-tag. Methods in Molecular Biology, 1355, 17. |
| 8. |
Induction of autophagy-dependent apoptosis in cancer cells through activation of ER stress: an uncovered anti-cancer mechanism by anti-alcoholism drug disulfiram.Am J Cancer Res. 2019 Jun 1;9(6):1266-1281. eCollection 2019. |
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 301-93531 | Phos-tag™ Biotin BTL-104 Phos-tag 生物素 |
10 mg | – |
| 308-97201 | Phos-tag™ Biotin BTL-111 1mM Aqueous Solution Phos-tag 生物素1mM水溶液 |
0.1 mL | – |
Phos-tag™ 丙烯酰胺 Phos-tag™ Acrylamide
- 产品特性
- 相关资料
- Q&A
- 参考文献
Phos-tag™ Acrylamide
SDS-PAGE 分离不同磷酸化水平的蛋白!
在不使用放射性同位素的情况下,利用 Phos-tag™ SDS-PAGE 即可分离不同条带中的磷酸化和非磷酸化蛋白。分离后的凝胶可用于 Western blotting 和质谱分析等后续实验。
Phos-tag™ SDS-PAGE 操作简单,只需在常规 SDS-PAGE 胶中加入 Phos-tag™ Acrylamide 和 Mn2+ 或者 Zn2+ 即可进行实验。在电泳过程中,磷酸化蛋白的磷酸基团与 Phos-tag™ 中的二价金属离子相结合,降低其迁移速度,从而可区分磷酸化与非磷酸化蛋白。
◆原理

◆优点、特色
● 采用 Phos-tag™ SDS-PAGE 可轻松分离磷酸化蛋白
无任何放射性元素及化学标记!
● 可检测不同磷酸化水平的磷酸化蛋白
无需任何磷酸化抗体!
● 适用于内源性蛋白的磷酸化分析!
◆案例、应用


【使用Phos-tag™ SDS-PAGE的磷酸化/非磷酸化蛋白比较】
我推荐使用Phos-tag ™ ——东京大学研究院医学研究科 小川觉之
Phos-tag ™ 是专为研究磷酸化蛋白而新开发出来的试剂。此产品使用方便,不但可用于体外实验,还能定量分析体内蛋白的磷酸化水平。Phos-tag ™ SDS-PAGE 可用于常规电泳实验,无需购买特殊设备,性价比高。传统蛋白磷酸化的研究需要特异的磷酸化抗体、RI 等其它试剂,操作复杂,花费大,且放射性元素会有安全隐患,而 Phos-tag ™ 的出现恰恰可以弥补这些缺点,为磷酸化蛋白研究提供新的方向。
磷酸化蛋白和非磷酸化蛋白利用Phos-tag ™ SDS-PAGE 的分离效果图

Lane 1 为非磷酸化蛋白,Lane 2-5 为磷酸化蛋白,各蛋白因磷酸化状态不同而条带迁移率也有所不同。
磷酸化/ 非磷酸化蛋白的数量比、磷酸化程度、磷酸化蛋白的丰度等都可根据条带迁移和条带浓度求得。
【资料提供】
日本东京大学研究生院医学系研究科
【二维电泳中的应用:分析 hnRNP K 磷酸化异构体】
小鼠巨噬细胞 J774.1 经 LPS 刺激后,裂解细胞,经过免疫沉淀法分离得到 hnRNP K 。在二维电泳中,一维是IPG 胶,二维是 Phos-tag ™ SDS-PAGE,可分离 hnRNP K 的异构体。利用质谱仪,可以确认不同的点代表不同的亚型或修饰蛋白。
二维电泳

同一个等电点的位置上,不同位点发生磷酸化都可以被区分开来(例: spots 6 vs. 8 and spots 4 vs. 7)
【参考文献】
Characterization of multiple alternative forms of heterogeneous nuclear ribonucleoprotein K by phosphate-affinity electrophoresis. Y. Kimura, K. Nagata, N Suzuki, R. Yokoyama, Y. Yamanaka, H. Kitamura, H. Hirano, and O. Ohara, Proteomics , Nov 2010; 10(21): 3884-95.
【结果提供】
横滨市立大学 生命纳米系统科学研究科 生物体超分子系统科学专业 木村弥生(Dr. Y. Kimura)、平野久(Dr. H. Hirano)理化学研究所 RCAI 小原收
【EGF 刺激前后MAPK 磷酸化水平的变化】
常规 SDS-PAGE 和Phos-tagTM SDS-PAGE 后迚行克疫印迹实验分析 EGF 刺激的 A431 细胞中 MAPK 磷酸化水平。

摘自 Kinoshita-Kikuta, E. et al., Mol.Cell. Proteomics. (2007)6: 356.
操作视频,请点击:
样品处理(TCA沉淀):http://labchem.fujifilm-wako.com.cn/resources/show/47.html
凝胶制备:http://labchem.fujifilm-wako.com.cn/resources/show/47.html
已公开的验证蛋白列表,请点击
Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构

特点:
与 -2 价磷酸根离子的亲和性和选择性高于其它阴离子
在 pH 5-8 的生理环境下生成稳定的复合物
◆原理
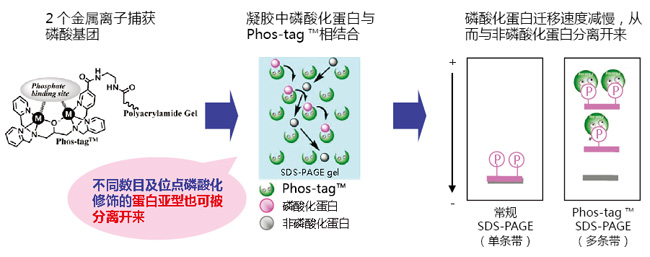
◆相关应用

◆相关产品
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离: SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离: 预制胶中含有50μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测: 代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化: 通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析: 用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™ 由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:

Phos-tag 第6版说明书

说明书
1. Phos-tag® Acrylamide的溶解
5 mmmol/ Phos-tag® 液体 (3v/v% 甲醇):
1) 10 mg Phos-tag® Acrylamide 里加入 0.1 mL 甲醇
2) 使用枪头搅拌混合直至完全溶解。
3) 加 3.2 mL 蒸馏水, 用枪头混匀。
2-8℃ 避光保存。不适合零度以下保存。建议保存时间6个月。
注意:避免溶解过程出现白色悬浮颗粒。
2. α-Casein, from Bovine Milk, Dephosphorylated(038-23221),阳性对照(含有磷酸化和非磷酸化α-Casein),如何使用?
用水或者上样 buffer 溶解。用水溶解后,冷冻保存。电泳条件:Phos-tag® 50 umol/L,分离胶浓度 10% 。
电流:30 mM,1小时。
3. 用Alkaline Phosphatase(for Biochemistry)(018-10693)进行磷酸化蛋白的去磷酸化反应体系。
37℃,过夜。# 10 mg/mL phosphorylated protein 50 μL
# 0.50 M Tris/HCl buffer (pH 9.0) containing 0.10 M MgCl2 10 μL
# Sterilized water 39 μL
# Alkaline phosphatase(018-10693). 0.3 unit / 1 μL 有一点需要注意:ALP 活性化使用 Mg 离子,相同的非磷酸化蛋白质用 ALP
处理的样品的条带和没有用ALP处理的样品的条带的位置不同。
4. Phos-tag® SDS-PAGE 实验没有成功分离磷酸化蛋白:
1) 使用 α-Casein, from Bovine Milk, Dephosphorylated(038-23221)作为阳性对照,确认实验条件和试剂均没有问题。
2) 可使用 Phos-tag® Biotin 检测样品中是否有磷酸化蛋白。确认有磷酸化蛋白后,再通过 Phos-tag® SDS-PAGE 进行分离鉴定。
3) 经质谱鉴定有表达磷酸化蛋白,建议增大样品的含量,可使用 Phos-tag® Agarose 进行磷酸化蛋白的富集。磷酸化蛋白含量过低,
会影响其分离效果。
4) 文献报道有表达磷酸化蛋白,或者同源蛋白有表达磷酸化蛋白的,建议用 Phos-tag® Biotin 先确认样品中是否有磷酸化蛋白。
5) 建议样品的 pH 值在7左右。酸性或者碱性条件下,Mn2+ -Phos-tag® 与磷酸化基团的特异性结合较差。
6) 避免样品中含有高浓度的还原剂,变性剂,表面活性剂等。β-巯基乙醇浓度不高于 0.2 M(或者5%)。
7) 进行 Phos-tag® SDS-PAGE 的最佳样品是纯化的蛋白。如果是细胞裂解液,体外激酶反应液,组织均浆液等,需要摸索最佳的分
离胶,Phos-tag® Acylamide 的浓度。建议 Phos-tag® Acrylamide 浓度从 50 μM 开始摸索。
5. Phos-tag® SDS-PAGE 凝胶用于 Western Blotting 实验的优化建议:
1) 可以检测的样品包括体外激酶反应体系,细胞裂解液,组织均浆液。
2) 每孔样品的上样量是 10~30 μg(请根据蛋白表达量进行调整)
3) 制备样品中含有的还原剂、变性剂、螯合剂、钒酸等会使电泳条带发生弯曲或者拖尾。通过 TCA 沉淀或渗析法降低杂质含量。
4) 建议样品的 pH 值在7左右。如果加入上样缓冲液后溶液显黄色或者橙色,加入 Tris 缓冲液调整 pH 值为7。
5) 目的蛋白分子量大于 60 kDa,分离胶的丙烯酰胺浓度为6%;目的蛋白分子量小于 60 kDa,分离胶的丙烯酰胺浓度为8%。
6) 如果样品中含有大量蛋白,比如细胞裂解液,组织均浆液,Phos-tag® Acylamide 浓度为 5~25 uM。
若目的蛋白浓度低,建议 Phos-tag® Acylamide 浓度为 100 uM。
7) Phos-tag® SDS-PAGE凝胶用于 Western Blotting 实验,湿法转膜建议:10 mM EDTA 的转移缓冲液处理凝胶 10 min,不含有
EDTA 的转移缓冲液处理凝胶 10 min。重复3次。强烈建议湿法转膜。
8) Phos-tag® SDS PAGE 半干法转膜建议:
i. 电泳后用含有 EDTA 的转移缓冲液处理凝胶,EDTA的浓度为 100 mM。100 mM EDTA 的转移缓冲液处理凝胶 10 min,不含有
EDTA的转移缓冲液处理凝胶 10 min。重复3次。
ii. 转膜的电流值提高2%~3%, 延长时间2成。
iii. 转膜的缓冲液加 SDS,加到大约 0.05~0.2%,转膜效率会提高。
9) 使用目的蛋白的非磷酸化抗体即可。比如检测各种肿瘤细胞系中 Src 激酶活性实验,用 Src 的非磷酸化抗体即可。
10) 富士胶片和光的 WIDE-VIEW™ Prestained Protein Siza MarkerIII(230-02461)可检测作为转膜效率,但是无法判断分子量。
11) 一般预染的蛋白 marker 在 Phos-tag® SDS-PAGE 中条带会弯曲,无法判断蛋白分子量。
12) 不能确认磷酸化蛋白和非磷酸化蛋白的分离,请进行常规的 SDS-PAGE,Western Blotting 实验。比对目的蛋白的迁移率。
13) 不能确认是因为蛋白发生磷酸化还是出现降解造成蛋白条带迁移,请进行常规的 SDS-PAGE 实验,确认不会出现条带迁移。
14) 目的蛋白磷酸化与非磷酸化分离效果不佳,使用 α-Casein、from Bovine Milk、Dephosphorylated(038-23221)作为阳性对照,
确认实验条件和试剂均没有问题。如果确认能够分离,调整分离胶,Phos-tag® Acylamide 的浓度。建议使用品质佳的 MnCl2
(139-00722)。
【参考文献】
· Conversion of graded phosphorylation into switch-like nuclear translocation via autoregulatory mechanisms in ERK signalling[J].Nature communications, 2016, 7,Shindo Y, Iwamoto K, Mouri K, et al.
· PTEN modulates EGFR late endocytic trafficking and degradation by dephosphorylating Rab7[J]. Nature communications, 2016, 7,Shinde S R, Maddika S.
· Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain[J]. Scientific Reports, 2016, 6: 31502,Kawasaki Y, Sakimura A, Park C M, et al.
· Plastid-nucleus communication involves calcium-modulated MAPK signalling[J]. Nature Communications, 2016, 7,Guo H, Feng P, Chi W, et al.
· Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation[J]. Nature communications, 2016, 7,Mitterer V, Murat G, Réty S, et al.
· Phos-tag analysis of Rab10 phosphorylation by LRRK2: a powerful assay for assessing kinase function and inhibitors[J]. Biochemical Journal, 2016: BCJ20160557,Ito G, Katsemonova K, Tonelli F, et al.
· Analysis of phosphorylation of the myosin targeting subunit of smooth muscle myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. The FASEB Journal, 2016, 30(1 Supplement): 1209.1-1209.1,Walsh M P, MacDonald J A, Sutherland C.
· Using Phos-Tag in Western Blotting Analysis to Evaluate Protein Phosphorylation[J]. Kidney Research: Experimental Protocols, 2016: 267-277,Horinouchi T, Terada K, Higashi T, et al.
· The Abundance of Nonphosphorylated Tau in Mouse and Human Tauopathy Brains Revealed by the Use of Phos-Tag Method[J]. The American journal of pathology, 2016, 186(2): 398-409,Kimura T, Hatsuta H, Masuda-Suzukake M, et al.
· Phos-tag SDS-PAGE resolves agonist-and isoform-specific activation patterns for PKD2 and PKD3 in cardiomyocytes and cardiac fibroblasts[J]. Journal of Molecular and Cellular Cardiology, 2016,Qiu W, Steinberg S F.
· Analysis of phosphorylation of the myosin-targeting subunit of myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. American Journal of Physiology-Cell Physiology, 2016, 310(8): C681-C691,Sutherland C, MacDonald J A, Walsh M P.
· Electrochemical biosensor for protein kinase A activity assay based on gold nanoparticles-carbon nanospheres, phos-tag-biotin and β-galactosidase[J]. Biosensors and Bioelectronics, 2016, 86: 508-515,Zhou Y, Yin H, Li X, et al.
· Validation of Cis and Trans Modes in Multistep Phosphotransfer Signaling of Bacterial Tripartite Sensor Kinases by Using Phos-Tag SDS-PAGE[J]. PloS one, 2016, 11(2): e0148294,Kinoshita-Kikuta E, Kinoshita E, Eguchi Y, et al.
· Phosphopeptide Detection with Biotin-Labeled Phos-tag[J]. Phospho-Proteomics: Methods and Protocols, 2016: 17-29,Kinoshita-Kikuta E, Kinoshita E, Koike T.
· A Phos‐tag SDS‐PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1[J]. Proteomics, 2016,Kinoshita E, Kinoshita‐Kikuta E, Kubota Y, et al.
· Difference gel electrophoresis of phosphoproteome: U.S. Patent Application 15/004,339[P]. 2016-1-22,Tao W A, Wang L.
· ERK1/2-induced phosphorylation of R-Ras GTPases stimulates their oncogenic potential[J]. Oncogene, 2016,Frémin C, Guégan J P, Plutoni C, et al.
· Microtubules Inhibit E-Cadherin Adhesive Activity by Maintaining Phosphorylated p120-Catenin in a Colon Carcinoma Cell Model[J]. PloS one, 2016, 11(2): e0148574,Maiden S L, Petrova Y I, Gumbiner B M.
· Serine 231 and 257 of Agamous-like 15 are phosphorylated in floral receptacles[J]. Plant Signaling & Behavior, 2016, 11(7): e1199314,Patharkar O R, Macken T A, Walker J C.
· A small molecule pyrazolo [3, 4-d] pyrimidinone inhibitor of zipper-interacting protein kinase suppresses calcium sensitization of vascular smooth muscle[J]. Molecular pharmacology, 2016, 89(1): 105-117,MacDonald J A, Sutherland C, Carlson D A, et al.
· The RNA polymerase II C-terminal domain phosphatase-like protein FIERY2/CPL1 interacts with eIF4AIII and is essential for nonsense-mediated mRNA decay in Arabidopsis[J]. The Plant Cell, 2016: TPC2015-00771-RA,Chen T, Qin T, Ding F, et al.
· Vasorelaxant Effect of 5′-Methylthioadenosine Obtained from Candida utilis Yeast Extract through the Suppression of Intracellular Ca2+ Concentration in Isolated Rat Aorta[J]. Journal of agricultural and food chemistry, 2016, 64(17): 3362-3370,Kumrungsee T, Akiyama S, Saiki T, et al.
· Inhibition of deubiquitinating activity of USP14 decreases tyrosine hydroxylase phosphorylated at Ser19 in PC12D cells[J]. Biochemical and biophysical research communications, 2016, 472(4): 598-602,Nakashima A, Ohnuma S, Kodani Y, et al.
· Actin Tyrosine-53-Phosphorylation in Neuronal Maturation and Synaptic Plasticity[J]. The Journal of Neuroscience, 2016, 36(19): 5299-5313,Bertling E, Englund J, Minkeviciene R, et al.
· AMPK-dependent phosphorylation of lipid droplet protein PLIN2 triggers its degradation by CMA[J]. Autophagy, 2016, 12(2): 432-438,Kaushik S, Cuervo A M.
· Myocardin-related transcription factor a and yes-associated protein exert dual control in G protein-coupled receptor-and RhoA-mediated transcriptional regulation and cell proliferation[J]. Molecular and cellular biology, 2016, 36(1): 39-49,Olivia M Y, Miyamoto S, Brown J H.
· Extensive phosphorylation of AMPA receptors in neurons[J]. Proceedings of the National Academy of Sciences, 2016, 113(33): E4920-E4927,Diering G H, Heo S, Hussain N K, et al.
· The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis[J]. The Plant Cell, 2016, 28(2): 557-567,Yamamoto Y, Negi J, Wang C, et al.
· The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of molecular and cellular cardiology, 2016, 90: 1-10,Kimura T E, Duggirala A, Smith M C, et al.
· Atg13 is essential for autophagy and cardiac development in mice[J]. Molecular and cellular biology, 2016, 36(4): 585-595,Kaizuka T, Mizushima N.
· The ChrSA and HrrSA two-component systems are required for transcriptional regulation of the hemA promoter in Corynebacterium diphtheriae[J]. Journal of Bacteriology, 2016: JB. 00339-16,Burgos J M, Schmitt M P.
· Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and Immunity, 2016, 84(7): 2086-2093,Zhu L, Olsen R J, Horstmann N, et al.
· Receptor for advanced glycation end products (RAGE) knockout reduces fetal dysmorphogenesis in murine diabetic pregnancy[J]. Reproductive Toxicology, 2016, 62: 62-70,Ejdesjö A, Brings S, Fleming T, et al.
· Aurora kinase-induced phosphorylation excludes transcription factor RUNX from the chromatin to facilitate proper mitotic progression[J]. Proceedings of the National Academy of Sciences, 2016, 113(23): 6490-6495,Chuang L S H, Khor J M, Lai S K, et al.
· Quantitative phosphoproteomics of protein kinase SnRK1 regulated protein phosphorylation in Arabidopsis under submergence[J]. Journal of experimental botany, 2016: erw107,Cho H Y, Wen T N, Wang Y T, et al.
· Temporal regulation of lipin activity diverged to account for differences in mitotic programs[J]. Current Biology, 2016, 26(2): 237-243,Makarova M, Gu Y, Chen J S, et al.
· Block of CDK1‐dependent polyadenosine elongation of Cyclin B mRNA in metaphase‐i‐arrested starfish oocytes is released by intracellular pH elevation upon spawning[J]. Molecular reproduction and development, 2016, 83(1): 79-87,Ochi H, Aoto S, Tachibana K, et al.
· Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1[J]. Cell reports, 2016, 15(9): 2050-2062,Rodriguez-Rodriguez J A, Moyano Y, Játiva S, et al.
· PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells[J]. Cancer medicine, 2016, 5(1): 74-87,Hu Z B, Liao X H, Xu Z Y, et al.
· Phosphorylated TDP-43 becomes resistant to cleavage by calpain: A regulatory role for phosphorylation in TDP-43 pathology of ALS/FTLD[J]. Neuroscience research, 2016, 107: 63-69,Yamashita T, Teramoto S, Kwak S.
· The Pch2 AAA+ ATPase promotes phosphorylation of the Hop1 meiotic checkpoint adaptor in response to synaptonemal complex defects[J]. Nucleic acids research, 2016: gkw506,Herruzo E, Ontoso D, González-Arranz S, et al.
· An optimized guanidination method for large‐scale proteomic studies[J]. Proteomics, 2016,Ye J, Zhang Y, Huang L, et al.
· Expression and purification of the kinase domain of PINK1 in Pichia pastoris[J]. Protein Expression and Purification, 2016,Wu D, Qu L, Fu Y, et al.
· BRI2 and BRI3 are functionally distinct phosphoproteins[J]. Cellular signalling, 2016, 28(1): 130-144,Martins F, Rebelo S, Santos M, et al.
· Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics[J]. Proteomics, 2016,Yang W, Jackson B, Zhang H.
· Regulation of Beclin 1 Protein Phosphorylation and Autophagy by Protein Phosphatase 2A (PP2A) and Death-associated Protein Kinase 3 (DAPK3)[J]. Journal of Biological Chemistry, 2016, 291(20): 10858-10866,Fujiwara N, Usui T, Ohama T, et al.
· Regulatory Implications of Structural Changes in Tyr201 of the Oxygen Sensor Protein FixL[J]. Biochemistry, 2016, 55(29): 4027-4035,Yamawaki T, Ishikawa H, Mizuno M, et al.
· Histone demethylase Jmjd3 regulates osteoblast apoptosis through targeting anti-apoptotic protein Bcl-2 and pro-apoptotic protein Bim[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(4): 650-659,Yang D, Okamura H, Teramachi J, et al.
· Analysis of Molecular Species Profiles of Ceramide-1-phosphate and Sphingomyelin Using MALDI-TOF Mass Spectrometry[J]. Lipids, 2016, 51(2): 263-270,Yamashita R, Tabata Y, Iga E, et al.
· Highly sensitive myosin phosphorylation analysis in the renal afferent arteriole[J]. Journal of Smooth Muscle Research, 2016, 52(0): 45-55,Takeya K.
· Functional dissection of the CroRS two-component system required for resistance to cell wall stressors in Enterococcus faecalis[J]. Journal of bacteriology, 2016, 198(8): 1326-1336,Kellogg S L, Kristich C J.
· Regulation of mitogen-activated protein kinase by protein kinase C and mitogen-activated protein kinase phosphatase-1 in vascular smooth muscle[J]. American Journal of Physiology-Cell Physiology, 2016, 310(11): C921-C930,Trappanese D M, Sivilich S, Ets H K, et al.
· ModProt: a database for integrating laboratory and literature data about protein post-translational modifications[J]. Journal of Electrophoresis, 2016, 60(1): 1-4,Kimura Y, Toda T, Hirano H.
· The C-ETS2-TFEB Axis Promotes Neuron Survival under Oxidative Stress by Regulating Lysosome Activity[J]. Oxidative medicine and cellular longevity, 2016,Ma S, Fang Z, Luo W, et al.
· Essential role of the PSI–LHCII supercomplex in photosystem acclimation to light and/or heat conditions by state transitions[J]. Photosynthesis Research, 2016: 1-10,Marutani Y, Yamauchi Y, Higashiyama M, et al.
· Identification of a redox-modulatory interaction between selenoprotein W and 14-3-3 protein[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(1): 10-18,Jeon Y H, Ko K Y, Lee J H, et al.
· Effects of hydrogen sulfide on the heme coordination structure and catalytic activity of the globin-coupled oxygen sensor AfGcHK[J]. BioMetals, 2016, 29(4): 715-729,Fojtikova V, Bartosova M, Man P, et al.
· Identification and functional analysis of phosphorylation in Newcastle disease virus phosphoprotein[J]. Archives of virology, 2016: 1-14,Qiu X, Zhan Y, Meng C, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports, 2016, 6: 54-62,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· Profiling DNA damage-induced phosphorylation in budding yeast reveals diverse signaling networks[J]. Proceedings of the National Academy of Sciences, 2016: 201602827,Zhou C, Elia A E H, Naylor M L, et al.
· Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system[J]. Nucleic Acids Research, 2016: gkw642,Brosse A, Korobeinikova A, Gottesman S, et al.
· Evolution of ZnII–Macrocyclic Polyamines to Biological Probes and Supramolecular Assembly[J]. Macrocyclic and Supramolecular Chemistry: How Izatt-Christensen Award Winners Shaped the Field, 2016: 415,Kimura E, Koike T, Aoki S.
· Phosphopeptide Enrichment Using Various Magnetic Nanocomposites: An Overview[J]. Phospho-Proteomics: Methods and Protocols, 2016: 193-209,Batalha Í L, Roque A C A.
· In vivo phosphorylation of a peptide tag for protein purification[J]. Biotechnology letters, 2016, 38(5): 767-772,Goux M, Fateh A, Defontaine A, et al.
· Regulation of cell reversal frequency in Myxococcus xanthus requires the balanced activity of CheY‐like domains in FrzE and FrzZ[J]. Molecular microbiology, 2016,Kaimer C, Zusman D R.
· Elevation of cortical serotonin transporter activity upon peripheral immune challenge is regulated independently of p38 mitogen‐activated protein kinase activation and transporter phosphorylation[J]. Journal of neurochemistry, 2016, 137(3): 423-435,Schwamborn R, Brown E, Haase J.
· The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression[J]. Molecular cell, 2016, 62(4): 532-545,Ewald J C, Kuehne A, Zamboni N, et al.
· Two Degradation Pathways of the p35 Cdk5 (Cyclin-dependent Kinase) Activation Subunit, Dependent and Independent of Ubiquitination[J]. Journal of Biological Chemistry, 2016, 291(9): 4649-4657,Takasugi T, Minegishi S, Asada A, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports. 2016,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· a high‐affinity LCO‐binding protein of Medicago truncatula, interacts with LYK3, a key symbiotic receptor[J]. FEBS letters, 2016, 590(10): 1477-1487,Fliegmann J, Jauneau A, Pichereaux C, et al. LYR3,
· Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability[J]. Molecular Cell, 2016,Spies J, Waizenegger A, Barton O, et al.
· PhostagTM-gel retardation and in situ thylakoid kinase assay for determination of chloroplast protein phosphorylation targets[J]. Endocytobiosis and Cell Research, 2016, 27(2): 62-70,Dytyuk Y, Flügge F, Czarnecki O, et al.
· Luteinizing Hormone Causes Phosphorylation and Activation of the cGMP Phosphodiesterase PDE5 in Rat Ovarian Follicles, Contributing, Together with PDE1 Activity, to the Resumption of Meiosis[J]. Biology of reproduction, 2016: biolreprod. 115.135897,Egbert J R, Uliasz T F, Shuhaibar L C, et al.
· Newby, AC, & Bond, M.(2016). The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of Molecular and Cellular Cardiology, 2016, 90: 1-10,Kimura-Wozniak T, Duggirala A, Smith M C, et al. G.
· Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels[J]. The FEBS Journal, 2016, 283(15): 2911-2928,Toume M, Tani M.
· Regulation of CsrB/C sRNA decay by EIIAGlc of the phosphoenolpyruvate: carbohydrate phosphotransferase system[J]. Molecular microbiology, 2016, 99(4): 627-639,Leng Y, Vakulskas C A, Zere T R, et al.
· The Late S-Phase Transcription Factor Hcm1 Is Regulated through Phosphorylation by the Cell Wall Integrity Checkpoint[J]. Molecular and cellular biology, 2016: MCB. 00952-15,Negishi T, Veis J, Hollenstein D, et al.
· Validation of chemical compound library screening for transcriptional co‐activator with PDZ‐binding motif inhibitors using GFP‐fused transcriptional co‐activator with PDZ‐binding motif[J]. Cancer science, 2016, 107(6): 791-802,Nagashima S, Maruyama J, Kawano S, et al.
· ULK1/2 Constitute a Bifurcate Node Controlling Glucose Metabolic Fluxes in Addition to Autophagy[J]. Molecular cell, 2016, 62(3): 359-370,Li T Y, Sun Y, Liang Y, et al.
· Spatiotemporal dynamics of Oct4 protein localization during preimplantation development in mice[J]. Reproduction, 2016: REP-16-0277,Fukuda A, Mitani A, Miyashita T, et al.
· The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress[J]. Parasites & Vectors, 2016, 9(1): 1,Pastor-Fernández I, Regidor-Cerrillo J, Álvarez-García G, et al.
· Interaction Analysis of a Two-Component System Using Nanodiscs[J]. PloS one, 2016, 11(2): e0149187,Hörnschemeyer P, Liss V, Heermann R, et al.
· Constitutive Activation of PINK1 Protein Leads to Proteasome-mediated and Non-apoptotic Cell Death Independently of Mitochondrial Autophagy[J]. Journal of Biological Chemistry, 2016, 291(31): 16162-16174,Akabane S, Matsuzaki K, Yamashita S, et al.
· p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261[J]. Molecular and cellular biology, 2016, 36(10): 1540-1554,Beenstock J, Melamed D, Mooshayef N, et al.
· Lysophosphatidylcholine acyltransferase 1 protects against cytotoxicity induced by polyunsaturated fatty acids[J]. The FASEB Journal, 2016, 30(5): 2027-2039,Akagi S, Kono N, Ariyama H, et al.
· Characterization of a herpes simplex virus 1 (HSV-1) chimera in which the Us3 protein kinase gene is replaced with the HSV-2 Us3 gene[J]. Journal of virology, 2016, 90(1): 457-473,Shindo K, Kato A, Koyanagi N, et al.
· Generation of phospho‐ubiquitin variants by orthogonal translation reveals codon skipping[J]. FEBS letters, 2016, 590(10): 1530-1542,George S, Aguirre J D, Spratt D E, et al.
· Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris[J]. PLoS Genet, 2016, 12(3): e1005922,Ma P, Mori T, Zhao C, et al.
· Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis[J]. FEMS Yeast Research, 2016, 16(6): fow060,Pérez J, Arcones I, Gómez A, et al.
· Alteration of Antiviral Signalling by Single Nucleotide Polymorphisms (SNPs) of Mitochondrial Antiviral Signalling Protein (MAVS)[J]. PloS one, 2016, 11(3): e0151173,Xing F, Matsumiya T, Hayakari R, et al.
· Arm-in-arm response regulator dimers promote intermolecular signal transduction[J]. Journal of bacteriology, 2016, 198(8): 1218-1229,Baker A W, Satyshur K A, Morales N M, et al.
· The lsh/ddm1 homolog mus-30 is required for genome stability, but not for dna methylation in neurospora crassa[J]. PLoS Genet, 2016, 12(1): e1005790,Basenko E Y, Kamei M, Ji L, et al.
· Fine tuning chloroplast movements through physical interactions between phototropins[J]. Journal of Experimental Botany, 2016: erw265,Sztatelman O, Łabuz J, Hermanowicz P, et al.
· Characterization of the Neospora caninum NcROP40 and NcROP2Fam-1 rhoptry proteins during the tachyzoite lytic cycle[J]. Parasitology, 2016, 143(01): 97-113,Pastor-Fernandez I, Regidor-Cerrillo J, Jimenez-Ruiz E, et al.
· Transcriptional Profile during Deoxycholate-Induced Sporulation in a Clostridium perfringens Isolate Causing Foodborne Illness[J]. Applied and environmental microbiology, 2016, 82(10): 2929-2942,Yasugi M, Okuzaki D, Kuwana R, et al.
· Timely Closure of the Prospore Membrane Requires SPS1 and SPO77 in Saccharomyces cerevisiae[J]. Genetics, 2016: genetics. 115.183939,Paulissen S M, Slubowski C J, Roesner J M, et al.
· DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach[J]. Nucleic Acids Research, 2016: gkw626,Wu K Z L, Wang G N, Fitzgerald J, et al.
· OVATE Family Protein 8 Positively Mediates Brassinosteroid Signaling through Interacting with the GSK3-like Kinase in Rice[J]. PLoS Genet, 2016, 12(6): e1006118,Yang C, Shen W, He Y, et al.
· Epithelial Sel1L is required for the maintenance of intestinal homeostasis[J]. Molecular biology of the cell, 2016, 27(3): 483-490, Sun S, Lourie R, Cohen S B, et al.
· Effect of Sodium Dodecyl Sulfate Concentration on Supramolecular Gel Electrophoresis[J]. ChemNanoMat, 2016,Tazawa S, Kobayashi K, Yamanaka M.
· Intergenic VNTR Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and immunity, 2016: IAI. 00258-16,Zhu L, Olsen R J, Horstmann N, et al.
· Ajuba Phosphorylation by CDK1 Promotes Cell Proliferation and Tumorigenesis[J]. Journal of Biological Chemistry, 2016: jbc. M116. 722751,Chen X, Stauffer S, Chen Y, et al.
· Editorial: International Plant Proteomics Organization (INPPO) World Congress 2014[J]. Frontiers in Plant Science, 2016, 7,Heazlewood J L, Jorrín-Novo J V, Agrawal G K, et al.
· Phosphoinositide kinase signaling controls ER-PM cross-talk[J]. Molecular biology of the cell, 2016, 27(7): 1170-1180,Omnus D J, Manford A G, Bader J M, et al.
· A multiple covalent crosslinked soft hydrogel for bioseparation[J]. Chemical Communications, 2016, 52(15): 3247-3250,Liu Z, Fan L, Xiao H, et al.
· Advances in crop proteomics: PTMs of proteins under abiotic stress[J]. Proteomics, 2016, 16(5): 847-865,Wu X, Gong F, Cao D, et al.
· Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae[J]. Molecular cell, 2016, 62(4): 546-557,Zhao G, Chen Y, Carey L, et al.
· Carbon Monoxide Gas Is Not Inert, but Global, in Its Consequences for Bacterial Gene Expression, Iron Acquisition, and Antibiotic Resistance[J]. Antioxidants & redox signaling, 2016,Wareham L K, Begg R, Jesse H E, et al.
· Two-layer regulation of PAQR3 on ATG14-linked class III PtdIns3K activation upon glucose starvation[J]. Autophagy, 2016: 1-2,Xu D, Wang Z, Chen Y.
· Regulation of sphingolipid biosynthesis by the morphogenesis checkpoint kinase Swe1[J]. Journal of Biological Chemistry, 2016, 291(5): 2524-2534,Chauhan N, Han G, Somashekarappa N, et al.
· PAX5 tyrosine phosphorylation by SYK co-operatively functions with its serine phosphorylation to cancel the PAX5-dependent repression of BLIMP1: A mechanism for antigen-triggered plasma cell differentiation[J]. Biochemical and biophysical research communications, 2016, 475(2): 176-181,Inagaki Y, Hayakawa F, Hirano D, et al.
· A Combined Computational and Genetic Approach Uncovers Network Interactions of the Cyanobacterial Circadian Clock[J]. Journal of Bacteriology, 2016: JB. 00235-16,Boyd J S, Cheng R R, Paddock M L, et al.
· HuR mediates motility of human bone marrow-derived mesenchymal stem cells triggered by sphingosine 1-phosphate in liver fibrosis[J]. Journal of Molecular Medicine, 2016: 1-14,Chang N, Ge J, Xiu L, et al.
· Combined replacement effects of human modified β-hexosaminidase B and GM2 activator protein on GM2 gangliosidoses fibroblasts[J]. Biochemistry and Biophysics Reports, 2016,Kitakaze K, Tasaki C, Tajima Y, et al.
· Roseotoxin B Improves Allergic Contact Dermatitis through a Unique Anti-inflammatory Mechanism Involving Excessive Activation of Autophagy in Activated T-Lymphocytes[J]. Journal of Investigative Dermatology, 2016,Wang X, Hu C, Wu X, et al.
References on Phos-tag™ Chemistry
· Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule, Rapid Communications of Mass Spectrometry, 17, 2075-2081 (2003), H. Takeda, A. Kawasaki, M. Takahashi, A. Yamada, and T. Koike
· Recognition of phosphate monoester dianion by an alkoxide-bridged dinuclear zinc (II) complex, Dalton Transactions, 1189-1193 (2004), E. Kinoshita, M. Takahashi, H. Takeda, M. Shiro, and T. Koike
· Quantitative analysis of lysophosphatidic acid by time-of-flight mass spectrometry using a phosphate capture molecule, Journal of Lipid Research, 45, 2145-2150 (2004), T. Tanaka, H. Tsutsui, K. Hirano, T. Koike, A. Tokumura, and K. Satouchi
· Production of 1,2-Didocosahexaenoyl Phosphatidylcholine by Bonito Muscle Lysophosphatidylcholine/Transacylase, Journal of Biochemistry,136, 477-483 (2004), K. Hirano, H. Matsui, T. Tanaka, F. Matsuura, K. Satouchi, and T. Koike
· Novel immobilized zinc(II) affinity chromatography for phosphopeptides and phosphorylated proteins, Journal of Separation Science, 28, 155-162 (2005), E. Kinoshita, A. Yamada, H. Takeda, E. Kinoshita-Kikuta, and T. Koike
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 304-93526 | Phos-tag Acrylamide AAL-107 5mM Aqueous Solution Phos-tag 丙烯酰胺5mM水溶液 |
0.3 mL | 蛋白研究 |
| 300-93523 | Phos-tag Acrylamide AAL-107 Phos-tag 丙烯酰胺 |
2 mg | 蛋白研究 |
| 304-93521 | Phos-tag Acrylamide AAL-107 Phos-tag 丙烯酰胺 |
10 mg | 蛋白研究 |
| 134-15302 | Manganese(II) Chloride Tetrahydrate氯化锰四水合物 | 25 g | for Molecular Biology |
WESTERNVIEW™ Detection Kits 无需猜测曝光时间:肉眼可见的信号显影
- 产品特性
- 相关资料
- Q&A
- 参考文献
无需猜测曝光时间:肉眼可见的信号显影![]()
WESTERNVIEW™ Detection Kits
用于Western blot的WESTERNVIEW™ 检测试剂盒,无需专用设备便可快速且清晰地显示转膜后的条带。
● 通过Enzo独有的多重酶技术,即可对低表达的标记物(如Caspase-7)进行高灵敏度检测
● 兼容PVDF和硝酸纤维素膜
● 简单且经济高效的方法
● 提供抗小鼠和抗兔检测试剂盒
请尝试使用Enzo的高灵敏度、易操作、信号持久的新型Western blot检测试剂盒。
◆清楚分辨多个目标

未使用以及使用Etoposide处理的PANC-1细胞。加入总细胞裂解物(20 µg),同时使用Tubulin和caspase抗体进行孵育,再用ENZ-KIT163进行显影。
◆无需昂贵设备即可快速获取清晰实验结果

来源于Jurkat细胞的总细胞裂解物。使用anti-tubulin进行检测,然后使用ENZ-KIT182进行显影,曝光时间1分钟。
◆可获取无背景的清晰条带

未使用以及使用12.5 μM的etoposide处理18 h的Jurkat细胞。裂解物上样量为12 μg/孔。
◆产品列表
|
产品编号 |
产品名称 |
包装 |
|
ENZ-KIT182-0001 |
WESTERNVIEW™Detection Kit (Anti-Mouse) |
1 Kit |
|
ENZ-KIT183-0001 |
WESTERNVIEW™Detection Kit (Anti-Rabbit) |
1 Kit |
|
ENZ-KIT163-0001 |
WESTERNVIEW Dual Detection Kit |
1 Kit |
◆相关产品
|
产品编号 |
产品名称 |
包装 |
|
ENZ-ACC131-0500 |
Prestained Protein Ladder (3.5 – 245 kDa) |
500 µL |
|
ENZ-KIT164-0001 |
PVDF Transfer Membrane (7 cm× 9 cm) |
10 EA |
|
ENZ-KIT165-0001 |
PVDF Transfer Membrane (10 cm×15 cm) |
10 EA |
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
Super Sep™ ACE Series 电泳聚丙烯酰胺凝胶
- 产品特性
- 相关资料
- Q&A
- 参考文献
电泳聚丙烯酰胺凝胶
Super Sep™ ACE Series
本产品具备高分离性能及稳定性,可长期保存。不仅可进行一般的 SDS-PAGE,也可进行 DNA 的电泳。凝胶不含 SDS,亦可用于 Native-PAGE。


戴着手套即可开封。无需剪刀。
◆利用不同浓度进行电泳

使用了预染 markerWIDE-VIEW™ Prestained Protein Size Marker Ⅲ(产品编号 230-02461)。
◆使用实例①
检测 His-tag 融合蛋白质(大肠杆菌表达)
利用 His-tag 的表达宿主(大肠杆菌,相当 50 mL 培养分量),对 His-tag 融合蛋白质的裂解条件进行实验。将各 45 μL 裂解样本与 15 μL 缓冲剂混合热处理后,15 μL/Lane 进行检测。
凝胶:Super Step™ ACE, 10-20%, 17 well(产品编号 198-15041)
样品缓冲剂:样品缓冲液(2ME+)(×4)(产品编号 191-13272)
电泳缓冲液:SDS-PAGE 缓冲液,pH8.5(产品编号 192-16801)
分子量marker:WIDE-VIEW™ Prestained Protein Size Marker Ⅲ(产品编号 230-02461)
CBB染色:QUICK-CBB(产品编号 299-50101)
镍琼脂糖凝胶:Ni-NTA Agarose HP(产品编号 145-09681)

◆使用实例②
检测 TARGET-tag 融合糖蛋白(HEK293N 细胞表达)
编码人源生理活性蛋白质基因的重组质粒载体,将其转染 HEK293N 细胞后,从细胞数 3×105 cells/mL 开始培养,0天后、3天后、7天后回收上清液。取上清液 20 μL/Lane,进行 SDS-PAGE,用银染及 Western 印迹方法检测出表达蛋白质。

| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 195-15171 | Super Sep™ ACE, 6%, 13well | 10包 | 电泳用 |
| 198-14941 | Super Sep™ ACE, 7.5%, 13well | 10包 | 电泳用 |
| 191-14931 | Super Sep™ ACE, 7.5%, 17well | 10包 | 电泳用 |
| 195-14951 | Super Sep™ ACE, 10%, 13well | 10包 | 电泳用 |
| 192-14961 | Super Sep™ ACE, 10%, 17well | 10包 | 电泳用 |
| 199-14971 | Super Sep™ ACE, 12.5%, 13well | 10包 | 电泳用 |
| 196-14981 | Super Sep™ ACE, 12.5%, 17well | 10包 | 电泳用 |
| 193-14991 | Super Sep™ ACE, 15%, 13well | 10包 | 电泳用 |
| 190-15001 | Super Sep™ ACE, 15%, 17well | 10包 | 电泳用 |
| 199-15191 | Super Sep™ ACE, 5-12%, 13well | 10包 | 电泳用 |
| 192-15201 | Super Sep™ ACE, 5-12%, 17well | 10包 | 电泳用 |
| 197-15011 | Super Sep™ ACE, 5-20%, 13well | 10包 | 电泳用 |
| 194-15021 | Super Sep™ ACE, 5-20%, 17well | 10包 | 电泳用 |
| 191-15031 | Super Sep™ ACE, 10-20%, 13well | 10包 | 电泳用 |
| 198-15041 | Super Sep™ ACE, 10-20%, 17well | 10包 | 电泳用 |
| 198-15301 | Super Sep™ ACE, 15-20%, 13well(甘氨酸凝胶) | 10包 | 电泳用 |
| 198-17361 | Super Sep™ ACE, 15-20%, 17well(甘氨酸凝胶) | 10包 | 电泳用 |
| 197-13291 | Super Sep™ 5~20%, 2D2维电泳凝胶。适用于7cm等电点电泳甘氨酸凝胶。 | 10包 | 电泳用 |
M65 EPIRAT™ ELISA PEVIVA 产品系列的新产品!
- 产品特性
- 相关资料
- Q&A
- 参考文献
PEVIVA 产品系列的新产品!![]()
M65 EPIRAT™ ELISA
◆M65 EpiRat™ ELISA

在 PEVIVA 产品线中,经过优化的新版 M65 EpiRat™ELISA 可特异性检测大鼠样本。此产品的推出,完成了整个 K18 系列产品的最后一块拼图,使 PEVIVA 产品系列可用于药物研发的所有阶段并具有更高的使用价值。

图1:药物研发中 PEVIVA产 品线的选择
随着 M65 EpiRat™ ELISA 这一新产品的到来,PEVIVA 产品线现在已经可以用于药物开发的各个阶段,从药物发现和临床前阶段直至药物的临床应用。
◆技术
M65 EpiRat™ ELISA 是基于与生物标志物角蛋白18(K18)结合的 M5 和 M6 抗体研发而成的。K18 的浓度反映了 K18 阳性细胞中包括凋亡和坏死在内的整体细胞死亡数量。当肝脏由于疾病或毒理作用而受损时,K18 的水平通常会升高。
◆毒理
M65 EpiRat™ ELISA 可用于毒理学研究。在全球传统毒理学测试指南中,通常使用啮齿类动物来预测人类健康的毒理结果,因此 M65 EpiRat™ 通过定量肝细胞中的死亡细胞来帮助评估肝毒性水平。
◆研究
在临床前研究中,M65 EpiRat™ ELISA 可用于分析药物治疗效果和疾病进展,例如新化合物在非酒精性脂肪肝(NAFL)和非酒精性脂肪性肝炎(NASH)中的作用。

图2
使用 M65 EpiRat™ ELISA 对未处理大鼠和使用不同剂量的扑热息痛(APAP)处理的大鼠的 11 份血浆样本进行分析。如图所示,相较于未处理大鼠,使用 1000 mg/kg APAP 和 1500 mg/kg APAP 处理的大鼠的 M65 EpiRat™ 值增加。每只大鼠分别在 0、24 和 48 h 后抽血。
◆特点
● 特异性测量大鼠 K18 阳性细胞中的总细胞死亡
● 定量大鼠血清和血浆样品中的 K18
● 简便的即用型产品,可根据实验情况进行拆分使用
◆产品列表
|
产品编号 |
产品名称 |
规格 |
|
10060 |
M65 EpiRat™ ELISA |
96 wells |
◆相关产品
|
产品编号 |
产品名称 |
规格 |
|
10040 |
M65 EpiDeath® ELISA |
96 wells |
|
10011 |
M30 Apoptosense® ELISA |
96 wells |
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
细胞凋亡 M30 CytoDeath™ ELISA试剂盒 M30 CytoDeath™ ELISA
- 产品特性
- 相关资料
- Q&A
- 参考文献
M30 CytoDeath™ 上皮细胞凋亡 ELISA 试剂盒![]()
M30 CytoDeath™ ELISA
◆原理
运用细胞培养物检测细胞凋亡。
M30 CytoDeath™ ELISA可用于细胞培养物的检测。可进行体外实验。
M30 CytoDeath™ ELISA可用于细胞凋亡诱导药物鉴定,包括时间进程动力学建立,剂量反应关系。也可用于大规模鉴定诱导细胞凋亡的 siRNA。该试剂盒可用于定量检测上皮细胞形成的多细胞球样体和器官培养体系的细胞凋亡(见 Herrmann et al., J Biomolecular Screening 13, 1, 2008)。
M30 CytoDeath™ ELISA 采用 M30 抗体(检测 K18 上 Asp396 caspase-cleavage 位点)和抗体 M6。该抗体组合不同于 M30 Apoptosense® ELISA 中的抗体,M30 CytoDeath™ ELISA 不能用于血清或者血浆。


◆案例・应用
在体外实验中定量检测上皮细胞的凋亡情况
用3D-细胞培养上清,人类器官(比如肝脏)、多细胞球样体(比如肿瘤干细胞)或组织(比如活体组织)培养进行毒理学研究;
可用于高通量筛选和鉴定促凋亡药物。
定量检测多细胞球样体
多细胞肿瘤球体是肿瘤3D微环境模型。作为抗癌药物的研究对象,球体优于单层细胞。
PEVIVA 与 Karolinska 研究所合作研发了用于多细胞球体药物筛选的试剂盒。M30 CytoDeath™ ELISA 给研究药物渗透和诱导细胞死亡的研究者提供了强有力的工具。
◆产品信息
M30 CytoDeath™ ELISA Kit
|
特异性 |
人、猴子、牛 |
|
样品类型 |
表达CK18的凋亡细胞和凋亡组织的细胞上清液和裂解液 |
|
灵敏度 |
60 U/L,Standard Z (0 U/L)+ 3 S.D. |
|
范围 |
250-3,000 U/L |
|
检测目标物 |
可溶性 caspase 酶解的 CK18(ccCK18/CK 18F/CK18Asp396-NE/M30) |
|
特点 |
检测范围广;样品制备容易;冷冻的细胞裂解液也可以进行检测;可用于自动化操作 |
| 稀释线性 | 用细胞培养基稀释,范围在80%-120%。 |
更多运用案例
 |
 |
|
PEVIVA |
PEVIVA肿瘤及肝病上皮细胞死亡检测试剂盒 |
相关文章:
PEVIVA M30 与 M65 试剂盒联用,判断细胞死亡形式
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 10900 | M30 CytoDeath™ ELISA M30 CytoDeath™ 上皮细胞凋亡ELISA试剂盒 |
96 wells |
细胞凋亡 M30 CytoDeath™ ELISA试剂盒 M30 CytoDeath™ ELISA
- 产品特性
- 相关资料
- Q&A
- 参考文献
细胞凋亡 M30 CytoDeath™ ELISA试剂盒
 |
 |
|
PEVIVA |
PEVIVA肿瘤及肝病上皮细胞死亡检测试剂盒 |
相关文章:
PEVIVA M30与M65试剂盒联用,判断细胞死亡形式
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 10700 | M30 CytoDEATH™ unlabeled | 200 tests | |
| 10750 | M30 CytoDEATH™ Biotin mAb | 200 tests | |
| 10800 | M30 CytoDEATH™ Fluorescein mAb | 200 tests | |
| 10830 | M30 CytoDEATH™ Orange mAb | 200 tests |
LabAssay™ Ammonia LabAssay™ 氨检测试剂盒
- 产品特性
- 相关资料
- Q&A
- 参考文献

LabAssay™ Ammonia
LabAssay™ 氨检测试剂盒
氨主要产生于肠道和肾脏。在生物体内产生的氨通过肝脏中的尿素循环通路转变成尿素,然后从尿液中排出。
本产品把亚硝基铁氰化钠作为催化剂,以基于苯酚、次氯酸的贝特洛反应的比色法为基础的氨检测试剂。
LabAssay™ Ammonia 是以人和小鼠等动物样品作为对象的生化学检测用试剂,可检测样品中的氨。
LabAssay™ 系列是研究用试剂。不可用于诊断。
检测样品:血液
检测物种:小鼠、大鼠、人
◆特点
● 少量样品即可进行检测
● 可使用微孔板检测多个样品
● 可检测小鼠等动物样品
◆检测原理
氨和苯酚在催化剂亚硝基铁氰化钠的作用下置换生成二氧二苯胺。在强碱存在下,利用与次氯酸钠反应生成呈蓝色的靛酚,通过测定吸光度,从而测定出样本中的氮氨浓度。

◆试剂盒组成
● 脱蛋白试剂——100 mL×1瓶
● 显色剂 A——50 mL×1瓶
● 显色剂 B——25 mL×1瓶
● 显色剂 C——50 mL×1瓶
● 氨标准溶液——15 mL×1瓶
● 标准溶液用稀释液——200 mL×1瓶
◆性能
● 标准曲线范围——100~400 μg/dL(μg/100 mL)
● 检测时间——约70分钟
● 所需样品量——70 μL
● 检测波长——630 nm
〈标准曲线示例〉

◆使用方法
【微孔板法】
添加经脱蛋白试剂处理的样品 70 μL、显色剂 A 70 μL、显色剂 B 35 μL、显色剂 C 70 μL 至微孔板,使用孔板混匀仪充分混合,在37°C下加热 20 min。再次充分混合,室温下静置 30 min。以空白为对照,检测样品和标准溶液的吸光度。
【试管法】
添加经脱蛋白试剂处理的样品 800 μL、显色剂 A 800 μL、显色剂 B 400 μL、显色剂 C 800 μL 至微管或试管,充分搅拌,并在37°C下加热 20 min。再次搅拌后,用水(20~25°C)冷却 30 min。以空白为对照,检测样品和标准溶液的吸光度。
※ 使用此方法在试管内检测时,试剂盒的使用次数为 60 次。
◆产品列表
|
产品编号 |
产品名称 |
规格 |
包装 |
|
633-51761 (旧编号:295-78901) |
LabAssay™ Ammonia LabAssay™ 氨检测试剂盒 |
细胞生物学用 |
700 tests |
◆相关产品
|
产品编号 |
产品名称 |
规格 |
包装 |
|
633-51021 (旧编号:291-58601) |
LabAssay™ ALP LabAssay™ 碱性磷酸酶检测试剂盒 |
细胞生物学用 |
500 tests |
|
635-50981 (旧编号:294-65801) |
LabAssay™ Cholestrol LabAssay™ 胆固醇检测试剂盒 |
细胞生物学用 |
500 tests |
|
636-51011 (旧编号:290-65901) |
LabAssay™ Creatinine LabAssay™ 肌酸检测试剂盒 |
细胞生物学用 |
500 tests |
|
638-50971 (旧编号:298-65701) |
LabAssay™ Glucose LabAssay™ 葡萄糖检测试剂盒 |
细胞生物学用 |
500 tests |
|
633-52001 (旧编号:294-63601) |
LabAssay™ NEFA LabAssay™ 游离脂肪酸检测试剂盒 |
细胞生物学用 |
500 tests |
|
639-51001 (旧编号:296-63801) |
LabAssay™ Phospholipid LabAssay™ 磷脂检测试剂盒 |
细胞生物学用 |
500 tests |
|
632-50991 (旧编号:290-63701) |
LabAssay™ Triglyceride LabAssay™ 甘油三酯检测试剂盒 |
细胞生物学用 |
350 tests |
点击了解LabAssay™全系列
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
生化检测用试剂盒 LabAssay™ 系列
- 产品特性
- 相关资料
- Q&A
- 参考文献

生化检测用试剂盒 LabAssay™ 系列
LabAssay™ 系列产品是以人、小鼠等动物样品为检测对象的生化检测用试剂盒。使用微孔板检测,只需少量样品,一次可进行多样品检测。
※ LabAssay™ 系列产品仅供实验研究用,不可用于临床诊断。
◆LabAssay™ 系列产品
|
产品编号 |
检测项目 |
检测对象 |
对象样品 |
标准曲线范围 |
样品量 |
检测时间 |
包装 |
|
633-51761 (旧编号:295-78901) |
Ammonia 氨中氮元素 |
小鼠、大鼠、人 |
血液 |
100~400 μg/dL |
70 μL |
约 70 min |
700 tests |
|
633-51021 (旧编号:291-58601) |
ALP 碱性磷酸酶 |
小鼠、大鼠、人 |
血清、血浆 |
0.0625~0.5 mmol/L |
20 μL |
约 20 min |
500 tests |
|
635-50981 (旧编号:294-65801) |
Cholesterol 胆固醇 |
小鼠、大鼠、人 |
血清、血浆 |
50~592.2 mg/dL |
2 μL |
约 10 min |
500 tests |
|
636-51011 (旧编号:290-65901) |
Creatinine 肌氨酸酐 |
小鼠、大鼠、人 |
血清、血浆 |
2.5~10 mg/dL |
50 μL |
约 40 min |
500 tests |
|
638-50971 (旧编号:298-65701) |
Glucose 葡萄糖 |
小鼠、大鼠、人 |
血清、血浆 |
50~500 mg/dL |
2 μL |
约 10 min |
500 tests |
|
633-52001 (旧编号:294-63601) |
NEFA |
小鼠、大鼠、人 |
血清、血浆 |
0.4~1.97 mEq/L |
4 μL |
约 10 min |
500 tests |
|
639-51001 (旧编号:296-63801) |
Phospholipid 磷脂 |
小鼠、大鼠、人 |
血清、血浆 |
75.0~596.1 mg/dL |
2 μL |
约 10 min |
500 tests |
|
632-50991 (旧编号:290-63701) |
Triglyceride 甘油三酯 |
小鼠、大鼠、人 |
血清、血浆 |
100~800 mg/dL |
2 μL |
约 10 min |
350 tests |
◆LabAssay ™ Ammonia
◆LabAssay™ 氨检测试剂盒
氨主要在肠内、肾脏中产生。
生物体内生成的氨,通过肝脏的尿素循环变成尿素,从尿液中排出体外。
检测原理(藤井・奥田法变法)
氨与苯酚、催化剂五氰基亚硝酰基铁(Ⅲ)钠反应 ,转化为二氧二苯胺。二氧二苯胺在碱性条件下与次氯酸钠反应,生成靛酚。
本产品通过靛酚呈现的蓝色测定吸光度,以此计算样品中的氨氮浓度。
试剂盒组成
1. 脱蛋白液——100 mL×1瓶
2. 显色液 A——50 mL×1瓶
3. 显色液 B——25 mL×1瓶
4. 显色液 C——50 mL×1瓶
5. 氨标准溶液——15 mL×1瓶
6. 标准溶液用稀释液——20 mL×1瓶
试剂盒性能
● 标准曲线范围:100~400 μg/dL(μg/100 mL)
● 检测时间:约 70 min
● 所需样品量:70 μL
● 测定波长:630 nm
标准曲线示例

使用文献(例)
1)Inokuma, K. et al. : Microb. Cell Fact., 17, 153 (2018).
|
产品编号 |
产品名称 |
规格 |
包装 |
|
633-51761 (旧编号:295-78901) |
LabAssay™ Ammonia LabAssay™ 氨检测试剂盒 |
细胞生物学 |
700 tests |
◆LabAssay™ ALP
◆LabAssay™ 碱性磷酸酶检测试剂盒
碱性磷酸酶(ALP)广泛分布于肝脏、骨骼和小肠等。尤其是在骨代谢的研究领域中,被用作骨形成标志物之一。
检测原理
本品为碱性磷酸酶检测试剂盒,以p–硝基苯磷酸为基质,通过测定在碱性磷酸酶作用下生成的 p–硝基苯酚含量,可测定碱性磷酸酶活性。
试剂盒内容
1. 基质片——10片
2. 基质溶解液——50 mL×1瓶
3. 终止液——50 mL×1瓶
4. 标准液——5 mL×1瓶
试剂盒性能
● 标准曲线范围:0.0625~0.5 mmol/L
● 检测时间:约 20 min
● 所需样品量:20 μL
● 测定波长:405 nm
标准曲线示例

参考文献(例)
1. Ito, S. et al. : J. Pharmacol. Exp. Ther., 333, 341 (2010). ※小鼠肾组织提取物
2. Matsuyama, A. et al. : Clin. Exp. Pharmacol. Physiol., 45, 75 (2018). ※MC3T3-E 细胞,C2C12 细胞
3. Chiba, T. et al. : J. Atheroscler. Thromb., 23, 1099 (2016). ※小鼠血浆
4. Kohno, Y. et al. : Stem Cell Res. Ther., 8, 115 (2017). ※骨膜间充质干细胞
5. Iwakura, T. et al. : J. Orthop. Res., 27, 208 (2009). ※人培养细胞提取物
6. Furuya, Y. et al. : J. Biol. Chem., 286, 37023 (2011). ※小鼠血清
7. Itoh, T. et al. : J. Biol. Chem., 284, 19272 (2009). ※小鼠培养细胞提取物
|
产品编号 |
产品名称 |
规格 |
包装 |
|
633-51021 (旧编号:291-58601) |
LabAssay™ ALP LabAssay™ 碱性磷酸酶检测试剂盒 |
细胞生物学 |
500 tests |
◆LabAssay ™ Cholesterol
◆LabAssay™ 胆固醇检测试剂盒
胆固醇是生物体细胞膜的主要成分,是许多动物类固醇合成的前体物质,被认为是造成动脉硬化等血管疾病的原因之一。
检测原理(胆固醇氧化酶・DAOS法)
胆固醇氧化酶与胆固醇反应生成的过氧化氢在过氧化物酶(HRP)的作用下,与N-乙基-N-(2-羟基-3-磺丙基)-3,5-二甲氧基苯胺钠(DAOS)、4 -氨基安替比林发生氧化缩合。
本试剂盒通过氧化缩合生成的蓝色色素检测吸光度,以此计算样品中总胆固醇的含量。
试剂盒组成
1. 缓冲液——150 mL×1瓶
2. 显色剂——150 mL 用×1瓶
3. 标准液——5 mL×1瓶
试剂盒性能
● 标准曲线范围:50~592.2 mg/dL(mg/100 mL)
● 测定时间:约 10 min
● 所需样品量:2 μL
● 测定波长:主波长 600 nm、副波长 700 nm
标准曲线示例

参考文献(例)
1. Kobayashi, Y. et al. : J. Pharmacogn. Nat. Prod., online (2015) doi: 10.4172/2472-0992.1000113 ※小鼠肝脏提取物
2. Gao, F. et al. : Evid. Based Complement. Alternat. Med., 2015, 801291 (2015). ※大鼠肝组织提取物
3. Yoshioka, H. and Onosaka, S. : Fundam. Toxicol. Sci., 3, 151 (2016). ※小鼠血浆
4. Fujii, N. et al. : Aging Cell, 16, 508 (2017). ※大鼠血浆
5. Ushio, M. et al. : Am. J. Physiol. Endocrinol. Metab., 305, E293 (2013). ※小鼠组织提取物
6. Kobayashi, Y. et al. : Biosci. Biotechnol. Biochem., 74, 2385 (2010). ※大鼠组织提取物
|
产品编号 |
产品名称 |
产品规格 |
|
635-50981 (旧编号:294-65801) |
LabAssay™ Cholesterol LabAssay™ 胆固醇检测试剂盒 |
500 tests |
◆LabAssay ™ Creatinine
◆LabAssay™ 肌氨酸酐检测试剂盒
肌酐是一种由肌肉和神经中的磷酸肌酸直接产生或通过肌酸脱水产生,经肾小球过滤后排出体外的代谢产物。
检测原理(Jaffé法)
本试剂盒通过测定碱性条件下肌酐与苦味酸反应产生的橙红色色素,以此计算样品中肌酐的含量。
试剂盒组成
1. 脱蛋白试剂——150 mL×1瓶
2. 苦味酸试剂——50 mL×1瓶
3. 75 mol/L 氢氧化钠溶液——50 mL×1瓶
4. 肌酐标准溶液——15 mL×1瓶
试剂盒性能
● 标准曲线范围:2.5~10 mg/dL(mg/100mL)
● 检测时间:约 40 min
● 所需样品量:50 μL
● 测定波长:520 nm
标准曲线示例

参考文献(例)
1. Kawamoto, T. et al. : Glycative Stress Res., 3, 15 (2016). ※人尿
2. Guan, Y. et al. : J. Pharmacol. Sci., 135, 81 (2017). ※小鼠尿液、血浆
3. Ito, K. et al. : Biol. Pharm. Bull., 38, 1169 (2015). ※大鼠尿液、血浆
4. Tahara, Y. et al. : Med. Chem. commun., 8, 415 (2017). ※小鼠血清
5. Nasrin, S. et al. : J. Pharmacol. Sci., 122, 270 (2013). ※大鼠尿
6. Toyama, K. et al. : Br. J. Pharmacol., 166, 1183 (2012). ※小鼠
|
产品编号 |
产品名称 |
规格 |
包装 |
|
636-51011 (旧编号:290-65901) |
LabAssay™ Creatine LabAssay™ 肌酸检测试剂盒 |
细胞生物学 |
500 tests |
◆LabAssay ™ Glucose
◆LabAssay™ 葡萄糖检测试剂盒
糖是生物的重要能量来源之一,受到生物体内多种因子的调节。
在溶液中,葡萄糖的α型和β型维持在一定的比例内。
检测原理(变旋酶/GOD法)
通过与变旋酶作用,将α-D-葡萄糖转化为β-D-葡萄糖。β-D-葡萄糖与葡萄糖氧化酶(GOD)反应生成的过氧化氢在过氧化物酶(POD)的作用下,与苯酚和4-氨基安替比林进行定量氧化缩合。
本试剂盒通过氧化缩合生成的红色色素测定吸光度,以此计算样本中的葡萄糖浓度。
试剂盒组成
1. 缓冲液——150 mL×1瓶
2. 显色剂——150 mL 用×1瓶
3. 葡萄糖标准溶液Ⅰ——3 mL×1瓶
4. 葡萄糖标准溶液Ⅱ——3 mL×1瓶
试剂盒性能
● 标准曲线范围:50~500 mg/dL(mg/100 mL)
● 检测时间:约 10 min
● 所需样品量:2 μL
● 测定波长:主波长 505 nm、副波长 600 nm
标准曲线示例

参考文献(例)
1. Yamashita, Y. et al. : Biosci. Biotechnol. Biochem., 77, 888 (2013). ※小鼠血浆
2. Narita, T. et al. : Exp. Gerontol., 104, 127 (2018). ※大鼠血浆
3. Yamasaki, M. et al. : Food Sci. Technol. Res., 21, 827 (2015). ※小鼠血清
4. Fan, Y. et al. : J. Biomed. Sci., 23, 56 (2016). ※小鼠血清
5. Wang, W. et al. : J. Renin Angiotensin Aldosterone Syst., 15, 384 (2014). ※小鼠血清
6. Yamashita, Y. et al. : J. Nutr. Sci., 1, e2 (2012). ※小鼠血浆
7. Maesako, M. et al. : Neurobiol. Aging., 33, 1011 (2012). ※小鼠血清
|
产品编号 |
产品名称 |
规格 |
包装 |
|
638-50971 (旧编号:298-65701) |
LabAssay™ Glucose LabAssay™ 葡萄糖检测试剂盒 |
细胞生物学 |
500 tests |
◆LabAssay™ NEFA
◆LabAssay™ 游离脂肪酸检测试剂盒
在血清中NEFA(Non-esterified Fatty Acid:游离脂肪酸)与白蛋白结合,被转运至外周组织,成为外周组织的重要能源来源。游离脂肪酸的浓度受到脂肪组织释放、外周组织消耗和肝脏摄取的调节。
检测原理(ACS・ACOD法)
游离脂肪酸在酰基辅酶A合成酶(ACS)的作用下生成酰基辅酶A(Acyl-CoA)。生成的酰基辅酶A与酰基辅酶A氧化酶(ACOD)反应生成过氧化氢,过氧化氢在过氧化物酶(POD)的作用下,与 3-甲基-N-乙基-N-(β-羟乙基)-苯胺(MEHA)和 4-氨基安替比林进行氧化缩合。
本试剂盒通过氧化缩合生成的蓝紫色色素测定吸光度,以此计算样品中的游离脂肪酸含量。
试剂盒组成
1. 显色剂 A——10 mL 用×4瓶
2. 显色剂 A 溶解液——45 mL×1瓶
3. 显色剂 B——20 mL 用×4瓶
4. 显色剂 B 溶解液——90 mL×1瓶
5. 标准溶液(油酸1 mEq/L)——7 mL×1瓶
※油酸 1 mEq=1 mmol
试剂盒性能
● 标准曲线范围:0.4~1.97 mEq/L
● 检测时间:约20 min
● 所需样品量:4 μL
● 测定波长:550 nm
标准曲线示例

参考文献(例)
1. Kobayashi, Y. et al. : J. Pharmacogn. Nat. Prod., online (2015) doi: 10.4172/2472-0992.1000113 ※小鼠肝组织提取物
2. Gao, F. et al. : Evid. Based Complement. Alternat. Med., 2015, 801291 (2015). ※大鼠肝组织提取物
3. Ogawa, K. et al. : Reprod. Med. Biol., online (2018). doi.org/10.1002/rmb2.12084 ※猪卵泡来源的卵泡液
4. Wang, F. et al. : J. Mol. Endocrinol., 52, 133 (2014). ※小鼠血浆
5. Chang, Y. C. et al. : EMBO Mol. Med., 5, 1165 (2013). ※小鼠血浆
6. Uchida, K. et al. : Exp. Anim., 58, 181 (2009). ※小鼠血清
|
产品编号 |
产品名称 |
规格 |
包装 |
|
633-52001 (旧编号:294-63601) |
LabAssay™ NEFA LabAssay™ 游离脂肪酸检测试剂盒 |
细胞生物学 |
500 tests |
◆LabAssay ™ Phospholipid
◆LabAssay™ 磷脂检测试剂盒
磷脂参与生物体内细胞膜的构成、脂肪的乳化/吸收以及血液凝固等重要功能。
检测原理(胆碱氧化酶・DAOS法)
磷脂被磷脂酶D水解后释放胆碱。生成的胆碱与在胆碱氧化酶下产生的过氧化氢在过氧化物酶(POD)的作用下,与N-乙基-N-(2-羟基-3-磺丙基)-3,5-二甲氧基苯胺钠(DAOS)和4-氨基安替比林进行氧化缩合。
本试剂盒通过氧化缩合生成的蓝色色素测定吸光度,以此计算样品中磷脂的含量。
试剂盒性能
1. 缓冲液——50 mL×3瓶
2. 显色剂——50 mL用×3瓶
3. 标准溶液——5 mL×1瓶
试剂盒性能
● 标准曲线范围:75~596.1 mg/dL
● 检测时间:约 10 min
● 所需样品量:2 μL
● 测定波长:主波长 505 nm、 副波长 600 nm
标准曲线示例

参考文献(例)
1. Xu, Q. et al. : Biosci. Biotechnol, Biochem., 77, 1390 (2013). ※小鼠肝脏粉碎液
2. Tatematsu, Y. et al. : Biol. Pharma. Bull., 41, 319 (2018). ※脂质体
3. Kuge, H. et al. : J. Biol. Chem., 289, 26783 (2014). ※脂质体
4. Kessler, E. C. et al. : J. Dairy. Sci., 97, 5481 (2014). ※牛血浆
5. Cheng, L. et al. : Transplantation, 90, 127 (2010). ※大鼠
|
产品编号 |
产品名称 |
规格 |
包装 |
|
639-51001 (旧编号:296-63801) |
LabAssay™ Phospholipid LabAssay™ 磷脂检测试剂盒 |
细胞生物学 |
500 tests |
◆LabAssay ™ Triglyceride
◆LabAssay™ 甘油三酯检测试剂盒
甘油三酯是由1分子甘油和3分子脂肪酸通过酯键结合而成的中性脂肪。甘油三酯、胆固醇、磷脂、游离脂肪酸和脂溶性维生素作为脂溶性物质存在于血液中。
检测原理
甘油三酯在脂蛋白脂肪酶和甘油激酶的作用下转化为 3-磷酸甘油。3-磷酸甘油与3-磷酸甘油氧化酶(GPO)反应生成的过氧化氢,在过氧化物酶(POD)的作用下,与N-乙基-N-(2-羟基-3-磺丙基)-3 ,5-二甲氧基苯胺钠(DAOS)和 4-氨基安替比林进行氧化缩合。
本试剂盒通过氧化缩合生成的蓝色色素测定吸光度,以此计算样品中甘油三酯的含量。
试剂盒组成
1. 缓冲液——105 mL×1瓶
2. 显色剂——105 mL用×1瓶
3. 标准液——4 mL×1瓶
试剂盒性能
● 标准曲线范围——100~888 mg/dL(mg/100mL)
● 检测时间——约 10 min
● 所需样品量——2 μL
● 测定波长——主波长 600 nm、 副波长 700 nm
标准曲线示例

参考文献(例)
1. Gao, F. et al. : Evid. Based Complement. Alternat. Med., 2015, 801291 (2015). ※大鼠肝组织提取物
2. Funakoshi, T. et al. : Biochem. Biophys. Rep., 13, 39 (2018). ※大鼠原代肌肉卫生细胞
3. Moser, V. A. and Pike, C. J. : eNeuro, 4, e0077-17 (2017). ※小鼠血浆
4. Fujii, N. et al. : Aging Cell, 16, 508 (2017). ※大鼠血浆
5. Wang, F. et al. : J. Mol. Endocrinol., 52, 133 (2014). ※大鼠血浆
6. Kato, H. et al. : J. Hepatol., 60, 1032 (2014). ※小鼠肝脏提取物
7. Fujimori, K. et al. : Prostaglandins. Other. lipid. Mediat., 94, 96 (2011). ※小鼠培养细胞提取物
|
产品编号 |
产品名称 |
规格 |
包装 |
|
632-50991 (旧编号:290-63701) |
LabAssay™ Triglyceride LabAssay™ 甘油三酯检测试剂盒 |
细胞生物学 |
350 tests |
◆肥胖·糖尿病相关试剂
|
产品编号 |
产品名称 |
检量线范围 |
规格 |
容量 |
|
299-75501 |
GLP-1ELISA试剂盒,高灵敏度 GLP-1 ELISA kit Wako,High Sensitive |
0.94~30 pmol/L(3.1~100 pg/mL) |
糖尿病研究用 |
96次用 |
|
291-73501 |
大鼠/小鼠PYY检测ELISA试剂盒 Mouse/Rat PYY ELISA Kit Wako |
0.15~12.5 ng/mL |
肥胖研究用 |
96次用 |
|
299-73801 |
小鼠活性GIP检测ELISA试剂盒 Mouse GIP(Active) ELISA Kit Wako |
7.8~500 pg/mL |
糖尿病研究用 |
96次用 |
|
297-57101 |
大鼠胰高血糖素ELISA试剂盒 Rat Glucagon ELISA Kit Wako |
41~10,000 pg/mL |
糖尿病研究用 |
96次用 |
|
292-60601 |
大鼠胰高血糖素样肽-2ELISA试剂盒 Rat GLP-2 ELISA Kit Wako |
0.137~100 ng/mL |
糖尿病研究用 |
96次用 |
|
295-57401 |
大鼠C肽ELISA试剂盒 Rat C-Peptide ELISA Kit Wako |
1.56~50 ng/mL |
糖尿病研究用 |
96次用 |
点击了解产品详情:
LabAssay™ 氨检测试剂盒
LabAssay™ 碱性磷酸酶检测试剂盒
LabAssay™ 胆固醇检测试剂盒
LabAssay™ 肌氨酸酐检测试剂盒
LabAssay™ 葡萄糖检测试剂盒
LabAssay™ NEFA检测试剂盒
LabAssay™ 磷脂检测试剂盒
LabAssay™ 甘油三酯检测试剂盒
◆LabAssay™ ALP(碱性磷酸酯检测试剂盒)
[1] Ito, S. et al.: J. Pharmacol. Exp. Ther., 333, 341 (2010). ※小鼠肾脏组织提取液
[2] Iwakura, T. et al.: J. Orthop. Res., 27, 208 (2009). ※人培养细胞提取液
[3] Furuya, Y. et al.: J. Biol. Chem., 286, 37023 (2011). ※小鼠血清
[4] Itoh, T. et al.: J. Biol. Chem., 284, 19272 (2009). ※小鼠培养细胞提取液
◆LabAssay™ Cholesterol(胆固醇检测试剂盒)
[1] Wu, S. Y. et al.: Acta. Pharmacol. Sin., 30, 202 (2009). ※小鼠血浆
[2] Uchida, K. et al.: Exp. Anim., 58, 181 (2009). ※小鼠组织提取液
[3] Ushio, M. et al.: Am. J. Physiol. Endocrinol. Metab., 305, E293 (2013). ※小鼠组织提取液
[4] Kobayashi, Y. et al.: Biosci. Biotechnol. Biochem., 74, 2385 (2010). ※大鼠至提取液
◆LabAssay™ Creatinine(肌氨酸酐检测试剂盒)
[1] Aranami, F. et al.: J. Med. Invest., 57, 95 (2010). ※小鼠血浆及尿液
[2] Toyama, K. et al.: Br. J. Pharmacol., 166, 1183 (2012). ※小鼠
[3] Matsumoto, A. et al.: Environ. Health Prev. Med., 15, 211 (2010). ※人尿
[4] Nasrin, S. et al.: J. Pharmacol. Sci., 122, 270 (2013). ※大鼠尿
LabAssay™ Glucose(葡萄糖检测试剂盒)
[1] Yamashita, Y. et al.: Biosci. Biotechnol.Biochem.,77, 888 (2013). ※小鼠血浆
[2] Yamashita, Y. et al.: J. Nutr. Sci., 1, e2 (2012). ※小鼠血浆
[3] Wang, W. et al.: J. Renin Angiotensin Aldosterone Syst., 15, 384 (2014). ※小鼠血清
[4] Maesako, M. et al.: Neurobiol. Aging.,33, 1011 (2012). ※小鼠血清
LabAssay™ NEFA(游离脂肪酸检测试剂盒)
[1] Wang, F. et al.: J. Mol. Endocrinol., 52, 133 (2014). ※小鼠血浆
[2] Kobayashi, Y. et al.: Biosci. Biotechnol.Biochem., 74, 2385 (2010). ※大鼠血浆
[3] Chang, Y. C. et al.: EMBO Mol. Med., 5, 1165 (2013). ※小鼠血浆
[4] Uchida, K. et al.: Exp. Anim., 58, 181 (2009). ※小鼠血清
◆LabAssay™ Phospholipid(磷脂检测试剂盒)
[1] Xu, Q. et al.: Biosci. Biotechnol, Biochem., 77, 1390 (2013). ※小鼠肝脏组织裂解液
[2] Kuge, H. et al.: J. Biol. Chem., 289, 26783 (2014). ※脂质体
[3] Kessler, E. C. et al.: J. Dairy. Sci., 97, 5481 (2014). ※牛
[4] Cheng, L. et al.: Transplantation, 90, 127 (2010). ※大鼠
◆LabAssay™ Triglyceride(甘油三酯检测试剂盒)
[1] Saito, M. et al.: Biosci. Biotechnol.Biochem.,76, 2049 (2012). ※小鼠血清
[2] Fujimori, K. et al.: Prostaglandins. Other.lipid. Mediat.,94, 96 (2011). ※小鼠培养细胞提取液
[3] Zhou, L. et al.: Am. J. Physiol. Renal. Physiol., 305, F1491 (2013). ※大鼠血浆
[4] Wu, S. Y. et al.: Acta. Pharmacol.Sin.,30, 202 (2009). ※大鼠血浆
[5] Wang, F. et al.: J. Mol. Endocrinol., 52, 133 (2014). ※大鼠血浆
[6] Kato, H. et al.: J. Hepatol., 60, 1032 (2014). ※小鼠肝脏提取液
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |