
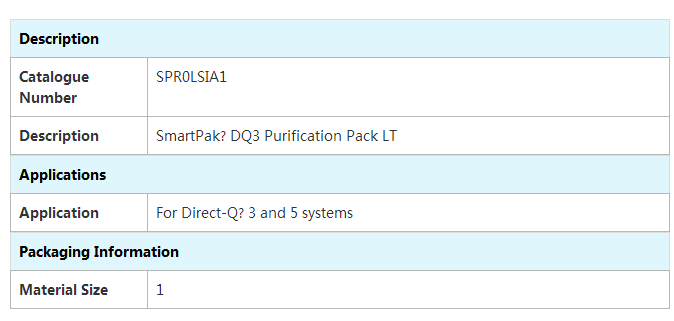
- 型号 SPR00SIA1
- 品牌 Merck Millipore密理博
- 【简单介绍】
默克millipore纯水柱SmartPak DQ3纯化柱 Direct-Q系统专为用水量较小的用户设计,可直接由自来水制备纯水和超纯水,适应实验室不同的应用需要。超纯水可用于色谱分离流动相制备,分光光度法空白和标准溶液的制备,光谱或其他分析技术,以及生化实验缓冲液的配制。
产品名称:纯化柱
规格型号:SmartPak DQ3
产品货号:SPR00SIA1
品牌|厂商:MERCK MILLIPORE|默克 密理博一体化Smartpak纯化柱便于简单且快速地进行更换
用活性炭预处理自来水,接着用反渗透法纯化水,然后再用离子交换树脂纯化水
一般应用
玻璃器皿清洗
增湿器、高压高温灭菌器、清洗机进水
Milli-Q? 超纯水系统的进水
直接生产 III 级实验室用水,产水流速可达 16L/h特性
预过滤柱装置包含三种纯化培养基
对反渗透膜的自行维护
带高回收回路的先进反渗透技术将水消耗充分降低 50%
稳定的产水流速
在对细菌敏感的应用中使用可选配的 UV 灯 可以获得*的水质
所有系统功能都可以通过袖珍键盘操作访问并显示在背光屏幕上
Direct-Q系统专为用水量较小的用户设计,可直接由自来水制备纯水和超纯水,适应实验室不同的应用需要。超纯水可用于色谱分离流动相制备,分光光度法空白和标准溶液的制备,光谱或其他分析技术,以及生化实验缓冲液的配制。纯水可用于常规玻璃器皿的洗涤和润洗。Direct-Q 3 UV系统专门配置了一个185/245nm双波长紫外灯,生产低于TOC水平的超纯水,尤其适合对有机物敏感的应用。创新的小体积远程取水器让您取水更便捷。
默克millipore纯水柱SmartPak DQ3纯化柱 SPR00SIA1
详情说明:
标签归档:纯化
CP2ALLRESCN-默克Millipore明澈A2纯化柱
- 型号 CP2ALLRESCN
- 品牌 Merck Millipore密理博
- 【简单介绍】
默克Millipore明澈A2纯化柱
说明:Q-GARD A2纯化柱
数量/包装:1
产品名称:Q-GARD 纯化柱
适用系统:MILLIPORE密理博实验室纯水超纯水系统明澈-D 24 UV
订货号:CP2ALLRESCN默克Millipore明澈A2纯化柱 CP2ALLRESCN
说明:Q-GARD A2纯化柱 数量/包装:1 产品名称:Q-GARD 纯化柱 适用系统:MILLIPORE密理博实验室纯水超纯水系统明澈-D 24 UV 订货号:CP2ALLRESCN
选配件:
储水水箱
可从Millipore水箱中选择容积为30L至350L不等的产品,以达到*纯水储量
纯水取水器:取水触手可及
从明澈-D系统可以选择纯水取水水枪,将储水水箱里的纯水通过取水水枪送到水池、清洗口、清洗机、老化箱等需要纯水的地方,从而极大的方便使用。
挂墙组件
将明澈-D安装在墙上或柜下,可节省空间。
保护装置
о漏水传感器–放置于地上,如果地板上有水,此传感器可使系统停止进水。
о液位传感器–将水箱液位信息传输给Milli-Q系统,以根据用户选择的等级启动或停止水纯化操作。当水箱空时,安全液面可阻止空气进入超纯水中。
о254nm UV灯:该UV灯为选配装置,从水箱入口上游处开始安装,可将即纯水中的细菌含量减少至原来的1/1000
终端过滤装置
可供选择的终端精制器,为您的研究带来Z适合的水质:
BioPak、Millipak精制器。
MERCK MILLIPORE默克密理博实验室纯水超纯水系统明澈-D 24 UV是您Z“忠实可靠的实验室纯水伙伴”,它能满足您对实验室纯水系统Z根本的需求。
默克Millipore明澈A2纯化柱 CP2ALLRESCN
SPR00S001-密理博RephiSolo S Pack RO3 纯化柱
- 型号 SPR00S001
- 品牌 Merck Millipore密理博
- 【简单介绍】
密理博RephiSolo S Pack RO3 纯化柱 SPR00S001
纯化方式:
活性碳
软化水介质
深层过滤介质
反渗透密理博RephiSolo S Pack RO3 纯化柱 SPR00S001
纯化方式:
活性碳
软化水介质
深层过滤介质
反渗透
QGARDT1X1-密理博millipore Q-Gard T1预纯化柱
- 型号 QGARDT1X1
- 品牌 Merck Millipore密理博
- 【简单介绍】
密理博millipore Q-Gard T1预纯化柱 QGARDT1X1
产品名称:Q-Gard T1预纯化柱产品货号:QGARDT1X1
品牌:密理博|Millipore
密理博货号: QGARDT1X1
纯化方式:
· 活性碳
· 离子交换
· LeFil™ 和 OrganeFil™ 配方填料
密理博millipore Q-Gard T1预纯化柱 QGARDT1X1
产品名称:Q-Gard T1预纯化柱
产品货号:QGARDT1X1
品牌:密理博|Millipore
密理博货号: QGARDT1X1
纯化方式:
· 活性碳
· 离子交换
· LeFil™ 和 OrganeFil™ 配方填料
密理博millipore Q-Gard T1预纯化柱 QGARDT1X1
订购信息:
QGARDT1X1 Q-GARD T1 PACK (1/PK)
QGARDT2X1 Q-GARD T2 PACK (1/PK)
QGARDT3X1 Q-GARD T3 PACK (1/PK)
QGARDTBX1 Q-GARD T BORON PACK (1/PK)
QGARDTL01 QGARD TL POLISHING PACK
QTUM0TIX1-密理博纯水柱Quantum TIX 纯化柱纯水机耗材
- 型号 QTUM0TIX1
- 品牌 Merck Millipore密理博
- 【简单介绍】
密理博纯水柱Quantum TIX 纯化柱纯水机耗材 QTUM0TIX1
Quantum TIX (Ionex 树脂) 纯化柱
商标名:Quantum
数量/包装:1
选择器标准:labwater
产品名称:Quantum TIX 柱说明:
Quantum TIX (Ionex 树脂) 纯化柱
商标名:
Quantum
数量/包装:
1
选择器标准:
labwater
产品名称:
Quantum TIX 柱
产品应用:
特别用于去除离子
RephiLe 货号: M1001MTIX
密理博货号: QTUM0TIX1
纯化方式:离子交换
LeFilTM 配方填料RephiSolo U Pack TIX 超纯化柱可为 Millipore 下述系统提供更换耗材:
Milli-Q Advantage A10 维护方案
Milli-Q Reference 维护方案
Milli-Q Integral 维护方案密理博纯水柱Quantum TIX 纯化柱纯水机耗材 QTUM0TIX1

SPR00SIA1-Millipore密理博DQ3纯化柱
- 型号 SPR00SIA1
- 品牌 Merck Millipore密理博
- 【简单介绍】
Millipore密理博DQ3纯化柱 SPR00SIA1
产品名称:纯化柱
规格型号:SmartPak DQ3
产品货号:SPR00SIA1
品牌|厂商:MERCK MILLIPORE|默克 密理博一体化Smartpak纯化柱便于简单且快速地进行更换
用活性炭预处理自来水,接着用反渗透法纯化水,然后再用离子交换树脂纯化水
Millipore密理博DQ3纯化柱 SPR00SIA1
产品名称:纯化柱
规格型号:SmartPak DQ3
产品货号:SPR00SIA1
品牌|厂商:MERCK MILLIPORE|默克 密理博一体化Smartpak纯化柱便于简单且快速地进行更换
用活性炭预处理自来水,接着用反渗透法纯化水,然后再用离子交换树脂纯化水
一般应用
玻璃器皿清洗
增湿器、高压高温灭菌器、清洗机进水
Milli-Q? 超纯水系统的进水
直接生产 III 级实验室用水,产水流速可达 16L/h特性
预过滤柱装置包含三种纯化培养基
对反渗透膜的自行维护
带高回收回路的先进反渗透技术将水消耗充分降低 50%
稳定的产水流速
在对细菌敏感的应用中使用可选配的 UV 灯 可以获得*的水质
所有系统功能都可以通过袖珍键盘操作访问并显示在背光屏幕上
Direct-Q系统专为用水量较小的用户设计,可直接由自来水制备纯水和超纯水,适应实验室不同的应用需要。超纯水可用于色谱分离流动相制备,分光光度法空白和标准溶液的制备,光谱或其他分析技术,以及生化实验缓冲液的配制。纯水可用于常规玻璃器皿的洗涤和润洗。Direct-Q 3 UV系统专门配置了一个185/245nm双波长紫外灯,生产低于TOC水平的超纯水,尤其适合对有机物敏感的应用。创新的小体积远程取水器让您取水更便捷。
Millipore密理博DQ3纯化柱 SPR00SIA1
产品详情:
WB120204-英国whatman FTA蛋白纯化试剂
- 型号 WB120204
- 品牌 GE whatman沃特曼
- 【简单介绍】
英国whatman FTA蛋白纯化试剂 WB120204
Whatman FTA纯化试剂 FTA REAGENT 500ML用于纯化保存在FTA卡上的核酸,确保得到高质量的DNA用于PCR或SNP分析,除去亚铁血红素,PCR抑制剂和其它潜在污染物,采用无毒,低盐的水溶液进行洗提FTA基因卡托盘,可同时放置2张FTA基因卡用于全自动液流分配/加样系统,托盘规格符合SBS标准订.英国whatman FTA蛋白纯化试剂 WB120204
Whatman FTA纯化试剂 FTA REAGENT 500ML用于纯化保存在FTA卡上的核酸,确保得到高质量的DNA用于PCR或SNP分析,除去亚铁血红素,PCR抑制剂和其它潜在污染物,采用无毒,低盐的水溶液进行洗提FTA基因卡托盘,可同时放置2张FTA基因卡用于全自动液流分配/加样系统,托盘规格符合SBS标准订.
简单介绍:
FTA纯化试剂&FTA基因卡托盘产品展台为您精选FTA纯化试剂&FTA基因卡托盘产品,FTA纯化试剂
●用于纯化保存在FTA卡上的核酸
●确保得到高质量的DNA用于PCR或SNP分析
●除去亚铁血红素,PCR抑制剂和其它潜在污染物
●采用无毒,低盐的水溶液进行洗提FTA基因卡托盘
●可同时放置2张FTA基因卡用于全自动液流分配/加样系统
●托盘规格符合SBS标准订
FTA纯化试剂&FTA基因卡托盘的详细介绍:
FTA纯化试剂&FTA基因卡托盘
产品介绍:
FTA纯化试剂
●用于纯化保存在FTA卡上的核酸
●确保得到高质量的DNA用于PCR或SNP分析
●除去亚铁血红素,PCR抑制剂和其它潜在污染物
●采用无毒,低盐的水溶液进行洗提
FTA基因卡托盘
●可同时放置2张FTA基因卡用于全自动液流分配/加样系统
●托盘规格符合SBS标准
GE whatman FTA微型卡用于DNA采集、纯化和分析的普通FTA卡英国whatman FTA蛋白纯化试剂 WB120204
订购信息:

PA标签系统 重组蛋白纯化检测用
- 产品特性
- 相关资料
- Q&A
- 参考文献

重组蛋白纯化检测用
PA标签系统


◆特点
● 通过树脂再生,降低运行成本
● 超高亲和性抗体,实现高回收率、高纯度纯化
● 在动物细胞表达系(HEK293T细胞、CHO细胞等)中,有效纯化蛋白
● PA标签系统是使用可识别肽标签(12个氨基酸残基)的单抗(NZ-1)。日本开发的亲和性纯化标签系统。
● NZ-1单抗对PA tag具有高亲和性和特异性,可应用于Western Blotting、细胞流式分析、免疫组化(ICC)。
通过免疫沉淀,可纯化PA标签融合蛋白以及树脂再生。
● PA标签系统,可一步纯化高纯度PA标签融合蛋白。
◆PA标签系统
Fujii, Y. et al., "PA tag: A versatile protein tagging system using a super high affi nity antibody against a dodecapeptide derived from human podoplanin.", Protein Exp. Purif., 95, 240-247(2014).
Fujii, Y. et al.
新型亲和标签系统“PA tag”的应用方法~蛋白的纯化及检测~
蛋白科学学会档案, 7, e075 (2014)
◆PA标签表达载体案例

|
载体 |
表达模式 |
宿主细胞 |
MCS启动子 |
|
pCAG |
瞬时表达 |
灵长类(人、猴)、犬细胞等粘着细胞 |
CAG |
和光还推出除上述抗性基因载体外的其他载体。(请点击参考。)
测序用引物序列、其他载体序列的相关详细信息,已附在产品现货说明书中。
另外可点击和光主页/TARGET标签,查看序列信息、Multi Cloning Site(MCS)信息。
◆用PA标签从培养上清中亲和提纯的案例

数据提供:大阪大学 蛋白研究所附属蛋白分析先进研究中心
分子创制学研究室 高木淳一老师,藤井勇树老师
◆Protocol目的蛋白的纯化
1.哺乳类动物细胞转染(HEK293T)
转染处于40-60%融合状态的细胞
培养基:D-MEM, 5%FBS
2.回收培养上清和吸附蛋白树脂
转染48-72小时后,回收约420 mL培养上清,加入8 mL抗PA标签抗体琼脂糖珠,混合2小时(2-10℃)
3.树脂回收
用50 mL的纯化柱。回收步骤2)中的琼脂糖珠
4.树脂洗涤
加入珠4倍量的TBS 缓冲液(20 mM Tris-HCl, 150 mM NaCl),清洗5次
5.PA标签肽从树脂中竞争性洗脱
加入经1×TBS溶液, pH 7.5溶解过的PA 标签肽(终浓度0.1 mg/mL),体积量需为珠的一倍,
然后分10次回收PA标签融合蛋白X
6.SDS-PAGE
样品量
c. sup:5 μL, flow through:5 μL, wash:5 μL,
肽洗脱(1-10):各10 μL
可纯化PA标签融合蛋白X且不依赖PA标签。→可实现极高纯度的亲和纯化。
◆通过PA tag降低背景的方法
|
|
M: Marker |

在20 mL的昆虫细胞溶解液中加入蛋白Z,使其浓度变为1.0 mg/L后,添加200 μL抗PA标签抗体琼脂糖珠,4℃温度条件下,搅拌3小时。
分别用不同浓度的TBS溶液(0.15 M、0.5 M、1.0 M NaCl)进行调整,用琼脂糖珠50倍量清洗。在回收的各珠中加入0.5 mL 0.1 M Glycine-HCl(pH 2.7),室温条件下平稳震动5 min。
离心后回收上清,用1 M Tris-HCl (pH 9.0)做中和处理。
SDS-PAGE后进行银染,比较去除污染蛋白之后,不同浓度下的结果。
◆使用案例:Western Blotting
|
地域革新领域 加藤 幸成老师,金子 美华老师 NZ-1显示出与天然抗体具有相同的高检测灵敏度。 |
Lane 1 一抗:抗IDH1抗体(RcMab-1), 1 μg/mL 二抗:抗大鼠IgG-HRP标签, 1000倍稀释 一抗:抗PA tag抗体, 1 μg/mL 二抗:抗大鼠IgG-HRP标签, 1000倍稀释 一抗:抗DYKDDDDK标签抗体, 3.5 μg/mL 二次抗体:抗小鼠IgG-HRP标签, 1000倍稀释 |

◆PA标签系列
|
产品名称 |
子类克隆号 |
适用实验 |
规格 |
产品编号 |
包装 |
|
单抗 |
|||||
|
抗PA标签, 大鼠单抗 Anti PA tag, Rat Monoclonal Antibody |
大鼠IgG2a |
IP, WB, FCM, ICC |
免疫化学用 |
016-25861 |
200 μL |
|
012-25863 |
1 mL |
||||
|
HRP标签单抗 |
|||||
|
抗PA标签, 大鼠单抗,过氧化酶结合 Anti PA tag, Rat Monoclonal Antibody, Peroxidase Conjugated |
大鼠IgG2a NZ-1 |
WB |
免疫化学用 |
015-25951 |
200 μL |
|
011-25953 |
1 mL |
||||
|
产品名称 |
子类克隆号 |
规格 |
产品编号 |
包装 |
|
|
抗体结合琼脂糖珠 |
|||||
|
抗PA标签抗体琼脂糖珠 Anti PA tag Antibody Beads |
大鼠IgG2a NZ-1 |
免疫化学用 |
012-25841 |
2 mL(Net 1 mL) |
|
|
018-25843 |
10 mL (Net 5 mL) |
||||
|
016-25844 |
50 mL (Net 25 mL) |
||||
|
识别N端、C端的PA标签 |
|||||
|
抗体结合磁珠 |
|||||
|
MagCaptureTM HP 抗PA标签抗体磁珠 MagCapture™ HP Anti PA tag Antibody Magnetic Beads |
大鼠IgG2a NZ-1 |
免疫化学用 |
137-18751 |
2 mL |
|
|
溶液组成:1×TBS(pH 7.4), 50% glycerol, 0.05 w/v% sodium azide |
|||||
|
产品名称 |
备注 |
规格 |
产品编号 |
包装 |
|
肽 |
||||
|
PA标签肽 PA tag Peptide |
含量(HPLC)95%以上 |
基因研究用 |
167-25501 |
5 mg |
|
163-25503 |
25 mg |
|||
|
清洗溶液 |
||||
|
PA标签清洗溶液 PA tag Washing Solution |
已过滤消毒 |
基因研究用 |
169-27261 |
50 mL |
|
产品名称 |
可选Marker |
产品编号 |
包装 |
|
|
大肠杆菌 |
动物细胞 |
|||
|
瞬时载体 |
||||
|
pCAG-Ble PA 标签-C pCAG-Ble PA tag-C |
Bleomycin |
161-26861 |
20 μg |
|
|
pCAG-Ble PA 标签-N 信号 增加型 pCAG-Ble PA tag-N Signal Plus |
Bleomycin |
168-26871 |
20 μg |
|
|
pCAG-Bsd PA 标签-C pCAG-Bsd PA tag-C |
Blasticidin S |
165-26881 |
20 μg |
|
|
pCAG-Bsd PA 标签-N 信号 增加型 pCAG-Bsd PA tag-N Signal Plus |
Blasticidin S |
162-26891 |
20 μg |
|
|
pCAG-Neo PA 标签-C pCAG-Hyg PA tag-C |
Hygromycin B |
165-26901 |
20 μg |
|
|
pCAG-Hyg PA 标签-N 信号 增加型 pCAG-Hyg PA tag-N Signal Plus |
Hygromycin B |
162-26911 |
20 μg |
|
|
pCAG-Neo PA 标签-C pCAG-Neo PA tag-C |
Kanamycin |
G418 |
169-26921 |
20 μg |
|
pCAG-Neo PA 标签-N 信号 增加型 pCAG-Neo PA tag-N Signal Plus |
Kanamycin |
G418 |
166-26931 |
20 μg |
◆相关产品
内参照
下表为Western Blot实验中参照用单抗一览表。
|
产品名称 |
规格 |
产品编号 |
包装 |
|
抗β肌动蛋白单抗 |
免疫化学用 |
013-24553 |
200 μg |
|
011-24554 |
1 mg |
||
|
017-24556 |
5 mg |
||
|
抗β肌动蛋白单抗,过氧化物酶标记 Anti β-Actin, Monoclonal Antibody, Peroxidase Conjugated |
免疫化学用 |
017-24573 |
200 μL |
|
抗α-微管蛋白, 单抗 Anti α-Tubulin, Monoclonal Antibody |
免疫化学用 |
011-25034 |
10 μg |
|
017-25031 |
200 μg |
||
|
013-25033 |
1 mg |
||
|
Anti β-Tubulin, Monoclonal Antibody |
免疫化学用 |
018-25044 |
10 μg |
|
014-25041 |
200 μg |
||
|
010-25043 |
1 mg |
||
|
Anti GAPDH, Monoclonal Antibody |
免疫化学用 |
016-25523 |
200 μg |
|
014-25524 |
1 mg |
||
|
010-25526 |
5 mg |
||
|
Anti GAPDH, Monoclonal Antibody, Peroxidase Conjugated |
免疫化学用 |
015-25473 |
200 μL |
蛋白酶抑制剂系列
|
产品名称 |
规格 |
产品编号 |
包装 |
|
蛋白酶抑制剂混合物试剂盒 I,无动物源成分(通用型) Protease Inhibitor Cocktail Set I, Animal-derived-free (for general use) (×100), Lyophilized |
通用型 |
165-26021 |
1 mL用 |
|
161-26023 |
1 mL用×5 |
||
|
蛋白酶抑制剂混合物试剂盒III, DMSO溶液(不含EDTA)(×100) |
动物细胞 |
163-26061 |
1 mL |
|
169-26063 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒V (无EDTA)(×100) |
2D电泳 |
162-26031 |
1 mL |
|
168-26033 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒 VI,DMSO溶液,用于植物(×100) |
植物细胞 |
164-26091 |
1 mL |
|
160-26093 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒VII,DMSO溶液,组氨酸标签蛋白(×100) |
His Tag 纯化※ |
167-26101 |
1 mL |
|
163-26103 |
1 mL×5 |
※ 本页面产品仅供研究用,研究以外不可使用。
亲和标签比较表
|
亲和标签 |
TARGET |
PA |
6xHis |
|
氨基酸序列 |
(YPGQ)5V |
GVAMPGAEDDVV |
HHHHHH |
|
残基数 |
21 |
12 |
6 |
|
分子量 (kDa) |
2.34 |
1.16 |
0.84 |
|
等电点(pI) |
5.52 |
3.49 |
7.21 |
|
配体 |
TARGET tag |
PA tag |
Ni, Co, Zn, Cu |
|
亲和珠回收 |
40% Propyleneglycol, |
3M MgCl2, MES(pH6.0) |
0.5M Imididazole(pH7.4) |
|
单抗边界强度 (KD(M)) |
1.0×10-8 |
4.9×10-10 |
Ni-NTA: |
|
单抗 Clone No. |
P20.1(Mouse) |
NZ-1(Rat) |
Merck:HIS. H8(Mouse) |
|
亲和力 |
++++ |
+++++ |
+++ |
|
提纯成本 |
+ |
+ |
+ |
|
多肽洗脱 |
+++ |
+++ |
Imidazole |
|
背景 (培养上清) |
+ |
+ |
+++++ |
|
背景 (细胞裂解物) |
+ |
+ |
++++ |
|
亲和标签 |
DYKDDDDK |
c-Myc |
HA |
GST |
|
氨基酸序列 |
DYKDDDDK |
EQKLISEEDL |
YPYDVPDYA |
– |
|
残基数 |
8 |
10 |
9 |
218 |
|
分子量 (kDa) |
1.01 |
1.20 |
1.10 |
26 |
|
等电点(pI) |
3.97 |
4.00 |
3.56 |
– |
|
配体 |
DYKDDDDK tag |
c-Myc tag |
HA tag |
GST |
|
亲和珠回收 |
Tris-HCl, |
Tris-HCl, |
Tris-HCl, |
Glutathione, |
|
单抗边界强度 (KD(M)) |
2.8×10-8 |
2.2×10-9 |
2.8×10-9 |
Glutathione: |
|
单抗 Clone No. |
M2(Mouse) |
9E10(Mouse) |
3F10(Mouse) |
Wako:5A7(Mouse) |
|
亲和力 |
++++ |
++++ |
++++ |
++++ |
|
提纯成本 |
++ |
++ |
++ |
+ |
|
多肽洗脱 |
+++ |
++ |
++ |
Glutathione |
|
背景 (培养上清) |
++++ |
+++ |
+++ |
++++ |
|
背景 (细胞裂解物) |
++++ |
++++ |
+++ |
++++ |
本比较表为Wako独家调查的数据。样品、检测方法和手法的不同,特异性也会有所变化,因此无法保证各个亲和标签的功能。
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
TARGET标签系统 重组蛋白纯化检测用
- 产品特性
- 相关资料
- Q&A
- 参考文献

重组蛋白纯化检测用
TARGET标签系统


◆特点
● 通过树脂再生,降低运作成本
● 温和洗脱、清洗,实现高纯度
● 在动物细胞表达系(HEK293T细胞、CHO细胞等)中有效纯化蛋白
TARGER标签系统使用的是可识别肽标签(21个氨基酸残基)的单抗(P20.1),是日本开发的亲和纯化标签系统。
TARGET标签系统实践证明,即使在传统系统都难以进行高纯度蛋白纯化的动物细胞表达系中,也可有效地进行亲和纯化。
◆TARGET标签系统
A rapid screening method for cell lines producing singly-tagged recombinant proteins using the“ TARGET tag”system. Tabata S. et al., J. Proteomics, 2010 Aug 5;73(9):1777-85.
◆TARGET 标签表达载体示意图

产品选择
|
载体 |
表达模式 |
宿主细胞 |
MCS启动子 |
|
pEBMulti |
非整合型稳定表达 |
灵长类(人、猴)、狗细胞等贴壁细胞 |
CAG |
|
pCAG |
瞬时表达 |
灵长类(人、猴)、狗细胞等贴壁细胞、大/小鼠等动物细胞 |
CAG |
富士胶片和光还推出除上述抗性基因载体外的其他载体。(请点击查看)
测序用引物序列、其他载体序列的相关详细信息,已附在产品随货说明书中。
另外可点击富士胶片和光主页/TARGET标签,查看序列信息、多克隆位点(MCS)信息。
|
TARGET标签融合蛋白的免疫沉淀
|
通过Transient 载体, 表达TARGET标签融合重组蛋白
|
|
培养上清(培养第7天) 每10 mL中,分别添加0.1 mL(net 0.05 mL)的抗体磁珠(①抗TARGET标签抗体琼脂糖珠、②抗DYKDDDDK标签抗体琼脂糖珠,其他公司产品),4℃条件下,进行2小时免疫沉淀。 清洗后,在回收的各珠中分别添加0.5 mL 0.2 mg/mL TARGET标签肽/TBS和0.2 mg/mL DYKDDDDK肽/TBS,4℃条件下,平稳震动0.5小时。离心后回收上清。 再分别在各珠中添加0.5 mL 0.1 M Glycine-HCl (pH 2.7),室温下,平稳震动5分钟。离心后,回收上清并用1M Tris-HCl (pH 9.0)中和。 上图为SDS-PAGE处理后的银染结果。 与传统DYKDDDDK tag相比,背景更低 →高纯度! |
M:M.W. Marker 1.仅转染载体的培养上清 2.转染了蛋白Y-TARGET 标签表达质粒的细胞培养上清 3.转染了TARGET标签-蛋白Y 表达质粒的细胞培养上清 也可使用市售的Transient载体 |
|
确认非整合型载体TARGET tag C端融合重组蛋白的表达 |
|
|
利用可编码表达人源生理活性蛋白基因的质粒转染 HEK293N 细胞后,从初期细胞数 3×105 cells/mL 开始培养,回收 0/3/7 日的培养上清。 用 20 μL/lane 的培养上清进行 SDS-PAGE 处理,通过银染和 Western blotting,检测出蛋白的表达。 |
|
|
可确认到与市面售卖的 Tag 系统有相同的表达量和检测灵敏度。 |
|




◆TARGET 标签系列
|
产品名称 |
子类 |
适用 实验 |
规格 |
产品编号 |
包装 |
|
|
单抗 |
||||||
|
抗TARGET标签,单抗 Anti TARGET tag,Monoclonal Antibody |
小鼠 IgG1 |
IP, WB |
免疫化学用 |
016-25481 |
200 μg |
|
|
012-25483 |
1 mg |
|||||
|
HRP标签单抗 |
||||||
|
抗 TARGET tag,单抗,过氧化酶结合 Anti TARGET tag, Monoclonal Antibody (P20.1) Peroxidase conjugated |
小鼠 IgG1 P20.1 |
WB |
免疫化学用 |
015-25571 |
200 μL |
|
|
011-25573 |
1 mL |
|||||
|
产品名称 |
子类克隆号 |
规格 |
产品编号 |
包装 |
|||||
|
抗体结合琼脂糖珠 |
|||||||||
|
抗 TARGET 标签抗体琼脂糖珠 Anti TARGET tag Antibody Beads |
小鼠 IgG1 P20.1 |
免疫化学用 |
018-25561 |
2 mL (Net 1 mL) |
|||||
|
014-25563 |
10 mL (Net 5 mL) |
||||||||
|
012-25564 |
50 mL (Net 25 mL) |
||||||||
|
识别 N 端、C 端的 TARGET 标签 使用载体:4% 琼脂糖 |
|||||||||
|
产品名称 |
备注 |
规格 |
产品编号 |
包装 |
|
肽 |
||||
|
TARGET标签肽 TARGET tag Peptide |
含量(HPLC)95%以上 |
基因研究用 |
200-19673 |
5 mg |
|
208-19674 |
25 mg |
|||
|
清洗液 |
||||
|
TARGET标签清洗液 TARGET tag Washing Solution |
已过滤消毒 |
基因研究用 |
208-19831 |
50 mL |
|
产品名称 |
可选 Marker |
产品编号 |
包装 |
||
|
大肠杆菌 |
动物细胞 |
||||
|
瞬时表达载体 |
|||||
|
C端插入TARGET标签的pCAG-Ble载体 pCAG-Ble TARGET tag-C |
博莱霉素 |
165-26381 |
20 μg |
||
|
N端插入TARGET标签的pCAG-Ble载体 pCAG-Ble TARGET tag-N |
博莱霉素 |
162-26391 |
20 μg |
||
|
C端插入TARGET标签的pCAG-Bsd载体 pCAG-Bsd TARGET tag-C |
杀稻瘟素S |
165-26401 |
20 μg |
||
|
N端插入TARGET标签的pCAG-Bsd载体 pCAG-Bsd TARGET tag-N |
杀稻瘟素S |
162-26411 |
20 μg |
||
|
C端插入TARGET标签的pCAG-Hyg载体 pCAG-Hyg TARGET tag-C |
潮霉素B |
169-26421 |
20 μg |
||
|
N端插入TARGET标签的pCAG-Hyg载体 pCAG-Hyg TARGET tag-N |
潮霉素B |
166-26431 |
20 μg |
||
|
C端插入TARGET标签的pCAG-Neo载体 pCAG-Neo TARGET tag-C |
卡那霉素 |
G418 |
163-26441 |
20 μg |
|
|
N端插入TARGET标签的pCAG-Neo载体 pCAG-Neo TARGET tag-N |
卡那霉素 |
G418 |
160-26451 |
20 μg |
|
|
游离载体 |
|||||
|
C段插入TARGET标签的pEBMulti-Ble载体 pEBMulti-Ble TARGET tag-C |
博来霉素 |
167-26461 |
20 μg |
||
|
N段插入TARGET标签的pEBMulti-Ble载体 pEBMulti-Ble TARGET tag-N |
博来霉素 |
164-26471 |
20 μg |
||
|
C段插入TARGET标签的pEBMulti-Bsd载体 pEBMulti-Bsd TARGET tag-C |
杀稻瘟素S |
161-26481 |
20 μg |
||
|
N段插入TARGET标签的pEBMultI-Bsd载体 pEBMulti-Bsd TARGET tag-N |
杀稻瘟素S |
168-26491 |
20 μg |
||
| C段插入TARGET标签的pEBMulti-Hyg载体
pEBMulti-Hyg TARGET tag-C |
潮霉素B |
161-26501 |
20 μg |
||
|
N段插入TARGET标签的pEBMulti-Hyg载体 pEBMulti-Hyg TARGET tag-N |
潮霉素B |
168-26511 |
20 μg |
||
|
C段插入TARGET标签的pEBMulti-Neo载体 pEBMulti-Neo TARGET tag-C |
卡那霉素 |
G418 |
165-26521 |
20 μg |
|
|
N段插入TARGET标签的pEBMulti-Neo载体 pEBMulti-Neo TARGET tag-N |
卡那霉素 |
G418 |
162-26531 |
20 μg |
|
|
C段插入TARGET标签的pEBMulti-Puro载体 pEBMulti-Puro TARGET tag-C |
氨苄青霉素 |
嘌呤霉素 |
169-26541 |
20 μg |
|
|
N段插入TARGET标签的pEBMulti-Puro载体 pEBMulti-Puro TARGET tag-N |
氨苄青霉素 |
嘌呤霉素 |
166-26551 |
20 μg |
|
◆相关产品
内参照
下表为 Western Blot 实验中参照用单抗一览表。
|
产品名称 |
规格 |
产品编号 |
包装 |
|
抗β肌动蛋白单抗 |
免疫化学用 |
013-24553 |
200 μg |
|
011-24554 |
1 mg |
||
|
017-24556 |
5 mg |
||
|
抗β肌动蛋白单抗,过氧化物酶标记 Anti β-Actin,Monoclonal Antibody,Peroxidase Conjugated |
免疫化学用 |
017-24573 |
200 μL |
|
抗α-微管蛋白,单抗 Anti α-Tubulin,Monoclonal Antibody |
免疫化学用 |
011-25034 |
10 μg |
|
017-25031 |
200 μg |
||
|
013-25033 |
1 mg |
||
|
Anti β-Tubulin,Monoclonal Antibody |
免疫化学用 |
018-25044 |
10 μg |
|
014-25041 |
200 μg |
||
|
010-25043 |
1 mg |
||
|
Anti GAPDH,Monoclonal Antibody |
免疫化学用 |
016-25523 |
200 μg |
|
014-25524 |
1 mg |
||
|
010-25526 |
5 mg |
||
|
Anti GAPDH,Monoclonal Antibody,Peroxidase Conjugated |
免疫化学用 |
015-25473 |
200 μL |
蛋白酶抑制剂系列
|
产品名称 |
规格 |
产品编号 |
包装 |
|
蛋白酶抑制剂混合物试剂盒 I,无动物源成分(通用型) Protease Inhibitor Cocktail Set I, Animal-derived-free (for general use) (×100), Lyophilized |
通用型 |
165-26021 |
1 mL用 |
|
161-26023 |
1 mL用×5 |
||
|
蛋白酶抑制剂混合物试剂盒III, DMSO溶液(不含EDTA)(×100) |
动物细胞 |
163-26061 |
1 mL |
|
169-26063 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒V (无EDTA)(×100) |
2D电泳 |
162-26031 |
1 mL |
|
168-26033 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒 VI,DMSO溶液,用于植物(×100) |
植物细胞 |
164-26091 |
1 mL |
|
160-26093 |
1 mL×5 |
||
|
蛋白酶抑制剂混合物试剂盒VII,DMSO溶液,组氨酸标签蛋白(×100) |
His Tag 纯化※ |
167-26101 |
1 mL |
|
163-26103 |
1 mL×5 |
亲和标签比较表
|
亲和标签 |
TARGET |
PA |
6xHis |
|
氨基酸序列 |
(YPGQ)5V |
GVAMPGAEDDVV |
HHHHHH |
|
残基数 |
21 |
12 |
6 |
|
分子量 (kDa) |
2.34 |
1.16 |
0.84 |
|
等电点(pI) |
5.52 |
3.49 |
7.21 |
|
配体 |
TARGET tag |
PA tag |
Ni, Co, Zn, Cu |
|
亲和珠回收 |
40% Propyleneglycol, |
3M MgCl2, MES(pH6.0) |
0.5M Imididazole(pH7.4) |
|
单抗边界强度 (KD(M)) |
1.0×10-8 |
4.9×10-10 |
Ni-NTA: |
|
单抗 Clone No. |
P20.1(Mouse) |
NZ-1(Rat) |
Merck:HIS. H8(Mouse) |
|
亲和力 |
++++ |
+++++ |
+++ |
|
提纯成本 |
+ |
+ |
+ |
|
多肽洗脱 |
+++ |
+++ |
Imidazole |
|
背景 (培养上清) |
+ |
+ |
+++++ |
|
背景 (细胞裂解物) |
+ |
+ |
++++ |
|
亲和标签 |
DYKDDDDK |
c-Myc |
HA |
GST |
|
氨基酸序列 |
DYKDDDDK |
EQKLISEEDL |
YPYDVPDYA |
– |
|
残基数 |
8 |
10 |
9 |
218 |
|
分子量 (kDa) |
1.01 |
1.20 |
1.10 |
26 |
|
等电点(pI) |
3.97 |
4.00 |
3.56 |
– |
|
配体 |
DYKDDDDK tag |
c-Myc tag |
HA tag |
GST |
|
亲和珠回收 |
Tris-HCl, |
Tris-HCl, |
Tris-HCl, |
Glutathione, |
|
单抗边界强度 (KD(M)) |
2.8×10-8 |
2.2×10-9 |
2.8×10-9 |
Glutathione: |
|
单抗 Clone No. |
M2(Mouse) |
9E10(Mouse) |
3F10(Mouse) |
Wako:5A7(Mouse) |
|
亲和力 |
++++ |
++++ |
++++ |
++++ |
|
提纯成本 |
++ |
++ |
++ |
+ |
|
多肽洗脱 |
+++ |
++ |
++ |
Glutathione |
|
背景 (培养上清) |
++++ |
+++ |
+++ |
++++ |
|
背景 (细胞裂解物) |
++++ |
++++ |
+++ |
++++ |
本比较表为Wako独家调查的数据。样品、检测方法和手法的不同,特异性也会有所变化,因此无法保证各个亲和标签的功能。
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
Phos-tag™ 琼脂糖 Phos-tag™ Agarose
- 产品特性
- 相关资料
- Q&A
- 参考文献
Phos-tag™ Agarose
亲和层析纯化磷酸化蛋白
填入色谱柱中使用。可分离、纯化、浓缩磷酸化蛋白。不使用界面活性剂、还原剂,可得到状态近似生物体内的磷酸化蛋白。
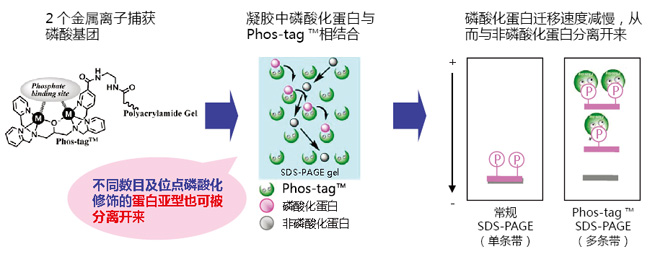
◆原理

◆优点、特色
● 缓冲液不含有界面活性剂、还原剂。
● 与亲和层析方法类似。
● 可在1小时内纯化磷酸化蛋白。
● Phos-tag™ Agarose 捕获结合到 Tyr、Thr、Ser、Asp、His 等氨基酸、糖类、脂类上的无机磷酸根和大量二价磷酸根。
● 可在生理条件下(pH7.5)捕捉蛋白。
● 纯化后的产物可用于 Co-IP 实验和其他蛋白活性实验。
◆案例、应用:
【使用例子:A431 裂解液中的磷酸化蛋白的纯化】
把Phos-tag™ 填充到柱里,再加上 A431 裂解液。
SYPRO Ruby 染色(左图)再使用 Anti-p Tyr 抗体进行免疫印迹(右图),检测出结果。
结果确认磷酸化蛋白浓缩在柱吸附层里。
M:分子量标记
Lane 1:未吸附层
Lane 2:吸附层
Lane 3:柱清洗层

Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot 检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™ Agarose)及质谱分析 MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构

◆特点
与 -2 价磷酸根离子的亲和性和选择性高于其它阴离子
在 pH 5-8 的生理环境下生成稳定的复合物
◆原理
◆相关应用

◆相关产品
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离:SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离:预制胶中含有 50 μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测:代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化:通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析:用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™ 由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:http://phos-tag.jp

Phos-tag 第6版说明书

Phos-tag系列 ver. 8

说明书
【参考文献】
· Conversion of graded phosphorylation into switch-like nuclear translocation via autoregulatory mechanisms in ERK signalling[J].Nature communications, 2016, 7,Shindo Y, Iwamoto K, Mouri K, et al.
· PTEN modulates EGFR late endocytic trafficking and degradation by dephosphorylating Rab7[J]. Nature communications, 2016, 7,Shinde S R, Maddika S.
· Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain[J]. Scientific Reports, 2016, 6: 31502,Kawasaki Y, Sakimura A, Park C M, et al.
· Plastid-nucleus communication involves calcium-modulated MAPK signalling[J]. Nature Communications, 2016, 7,Guo H, Feng P, Chi W, et al.
· Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation[J]. Nature communications, 2016, 7,Mitterer V, Murat G, Réty S, et al.
· Phos-tag analysis of Rab10 phosphorylation by LRRK2: a powerful assay for assessing kinase function and inhibitors[J]. Biochemical Journal, 2016: BCJ20160557,Ito G, Katsemonova K, Tonelli F, et al.
· Analysis of phosphorylation of the myosin targeting subunit of smooth muscle myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. The FASEB Journal, 2016, 30(1 Supplement): 1209.1-1209.1,Walsh M P, MacDonald J A, Sutherland C.
· Using Phos-Tag in Western Blotting Analysis to Evaluate Protein Phosphorylation[J]. Kidney Research: Experimental Protocols, 2016: 267-277,Horinouchi T, Terada K, Higashi T, et al.
· The Abundance of Nonphosphorylated Tau in Mouse and Human Tauopathy Brains Revealed by the Use of Phos-Tag Method[J]. The American journal of pathology, 2016, 186(2): 398-409,Kimura T, Hatsuta H, Masuda-Suzukake M, et al.
· Phos-tag SDS-PAGE resolves agonist-and isoform-specific activation patterns for PKD2 and PKD3 in cardiomyocytes and cardiac fibroblasts[J]. Journal of Molecular and Cellular Cardiology, 2016,Qiu W, Steinberg S F.
· Analysis of phosphorylation of the myosin-targeting subunit of myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. American Journal of Physiology-Cell Physiology, 2016, 310(8): C681-C691,Sutherland C, MacDonald J A, Walsh M P.
· Electrochemical biosensor for protein kinase A activity assay based on gold nanoparticles-carbon nanospheres, phos-tag-biotin and β-galactosidase[J]. Biosensors and Bioelectronics, 2016, 86: 508-515,Zhou Y, Yin H, Li X, et al.
· Validation of Cis and Trans Modes in Multistep Phosphotransfer Signaling of Bacterial Tripartite Sensor Kinases by Using Phos-Tag SDS-PAGE[J]. PloS one, 2016, 11(2): e0148294,Kinoshita-Kikuta E, Kinoshita E, Eguchi Y, et al.
· Phosphopeptide Detection with Biotin-Labeled Phos-tag[J]. Phospho-Proteomics: Methods and Protocols, 2016: 17-29,Kinoshita-Kikuta E, Kinoshita E, Koike T.
· A Phos‐tag SDS‐PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1[J]. Proteomics, 2016,Kinoshita E, Kinoshita‐Kikuta E, Kubota Y, et al.
· Difference gel electrophoresis of phosphoproteome: U.S. Patent Application 15/004,339[P]. 2016-1-22,Tao W A, Wang L.
· ERK1/2-induced phosphorylation of R-Ras GTPases stimulates their oncogenic potential[J]. Oncogene, 2016,Frémin C, Guégan J P, Plutoni C, et al.
· Microtubules Inhibit E-Cadherin Adhesive Activity by Maintaining Phosphorylated p120-Catenin in a Colon Carcinoma Cell Model[J]. PloS one, 2016, 11(2): e0148574,Maiden S L, Petrova Y I, Gumbiner B M.
· Serine 231 and 257 of Agamous-like 15 are phosphorylated in floral receptacles[J]. Plant Signaling & Behavior, 2016, 11(7): e1199314,Patharkar O R, Macken T A, Walker J C.
· A small molecule pyrazolo [3, 4-d] pyrimidinone inhibitor of zipper-interacting protein kinase suppresses calcium sensitization of vascular smooth muscle[J]. Molecular pharmacology, 2016, 89(1): 105-117,MacDonald J A, Sutherland C, Carlson D A, et al.
· The RNA polymerase II C-terminal domain phosphatase-like protein FIERY2/CPL1 interacts with eIF4AIII and is essential for nonsense-mediated mRNA decay in Arabidopsis[J]. The Plant Cell, 2016: TPC2015-00771-RA,Chen T, Qin T, Ding F, et al.
· Vasorelaxant Effect of 5′-Methylthioadenosine Obtained from Candida utilis Yeast Extract through the Suppression of Intracellular Ca2+ Concentration in Isolated Rat Aorta[J]. Journal of agricultural and food chemistry, 2016, 64(17): 3362-3370,Kumrungsee T, Akiyama S, Saiki T, et al.
· Inhibition of deubiquitinating activity of USP14 decreases tyrosine hydroxylase phosphorylated at Ser19 in PC12D cells[J]. Biochemical and biophysical research communications, 2016, 472(4): 598-602,Nakashima A, Ohnuma S, Kodani Y, et al.
· Actin Tyrosine-53-Phosphorylation in Neuronal Maturation and Synaptic Plasticity[J]. The Journal of Neuroscience, 2016, 36(19): 5299-5313,Bertling E, Englund J, Minkeviciene R, et al.
· AMPK-dependent phosphorylation of lipid droplet protein PLIN2 triggers its degradation by CMA[J]. Autophagy, 2016, 12(2): 432-438,Kaushik S, Cuervo A M.
· Myocardin-related transcription factor a and yes-associated protein exert dual control in G protein-coupled receptor-and RhoA-mediated transcriptional regulation and cell proliferation[J]. Molecular and cellular biology, 2016, 36(1): 39-49,Olivia M Y, Miyamoto S, Brown J H.
· Extensive phosphorylation of AMPA receptors in neurons[J]. Proceedings of the National Academy of Sciences, 2016, 113(33): E4920-E4927,Diering G H, Heo S, Hussain N K, et al.
· The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis[J]. The Plant Cell, 2016, 28(2): 557-567,Yamamoto Y, Negi J, Wang C, et al.
· The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of molecular and cellular cardiology, 2016, 90: 1-10,Kimura T E, Duggirala A, Smith M C, et al.
· Atg13 is essential for autophagy and cardiac development in mice[J]. Molecular and cellular biology, 2016, 36(4): 585-595,Kaizuka T, Mizushima N.
· The ChrSA and HrrSA two-component systems are required for transcriptional regulation of the hemA promoter in Corynebacterium diphtheriae[J]. Journal of Bacteriology, 2016: JB. 00339-16,Burgos J M, Schmitt M P.
· Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and Immunity, 2016, 84(7): 2086-2093,Zhu L, Olsen R J, Horstmann N, et al.
· Receptor for advanced glycation end products (RAGE) knockout reduces fetal dysmorphogenesis in murine diabetic pregnancy[J]. Reproductive Toxicology, 2016, 62: 62-70,Ejdesjö A, Brings S, Fleming T, et al.
· Aurora kinase-induced phosphorylation excludes transcription factor RUNX from the chromatin to facilitate proper mitotic progression[J]. Proceedings of the National Academy of Sciences, 2016, 113(23): 6490-6495,Chuang L S H, Khor J M, Lai S K, et al.
· Quantitative phosphoproteomics of protein kinase SnRK1 regulated protein phosphorylation in Arabidopsis under submergence[J]. Journal of experimental botany, 2016: erw107,Cho H Y, Wen T N, Wang Y T, et al.
· Temporal regulation of lipin activity diverged to account for differences in mitotic programs[J]. Current Biology, 2016, 26(2): 237-243,Makarova M, Gu Y, Chen J S, et al.
· Block of CDK1‐dependent polyadenosine elongation of Cyclin B mRNA in metaphase‐i‐arrested starfish oocytes is released by intracellular pH elevation upon spawning[J]. Molecular reproduction and development, 2016, 83(1): 79-87,Ochi H, Aoto S, Tachibana K, et al.
· Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1[J]. Cell reports, 2016, 15(9): 2050-2062,Rodriguez-Rodriguez J A, Moyano Y, Játiva S, et al.
· PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells[J]. Cancer medicine, 2016, 5(1): 74-87,Hu Z B, Liao X H, Xu Z Y, et al.
· Phosphorylated TDP-43 becomes resistant to cleavage by calpain: A regulatory role for phosphorylation in TDP-43 pathology of ALS/FTLD[J]. Neuroscience research, 2016, 107: 63-69,Yamashita T, Teramoto S, Kwak S.
· The Pch2 AAA+ ATPase promotes phosphorylation of the Hop1 meiotic checkpoint adaptor in response to synaptonemal complex defects[J]. Nucleic acids research, 2016: gkw506,Herruzo E, Ontoso D, González-Arranz S, et al.
· An optimized guanidination method for large‐scale proteomic studies[J]. Proteomics, 2016,Ye J, Zhang Y, Huang L, et al.
· Expression and purification of the kinase domain of PINK1 in Pichia pastoris[J]. Protein Expression and Purification, 2016,Wu D, Qu L, Fu Y, et al.
· BRI2 and BRI3 are functionally distinct phosphoproteins[J]. Cellular signalling, 2016, 28(1): 130-144,Martins F, Rebelo S, Santos M, et al.
· Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics[J]. Proteomics, 2016,Yang W, Jackson B, Zhang H.
· Regulation of Beclin 1 Protein Phosphorylation and Autophagy by Protein Phosphatase 2A (PP2A) and Death-associated Protein Kinase 3 (DAPK3)[J]. Journal of Biological Chemistry, 2016, 291(20): 10858-10866,Fujiwara N, Usui T, Ohama T, et al.
· Regulatory Implications of Structural Changes in Tyr201 of the Oxygen Sensor Protein FixL[J]. Biochemistry, 2016, 55(29): 4027-4035,Yamawaki T, Ishikawa H, Mizuno M, et al.
· Histone demethylase Jmjd3 regulates osteoblast apoptosis through targeting anti-apoptotic protein Bcl-2 and pro-apoptotic protein Bim[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(4): 650-659,Yang D, Okamura H, Teramachi J, et al.
· Analysis of Molecular Species Profiles of Ceramide-1-phosphate and Sphingomyelin Using MALDI-TOF Mass Spectrometry[J]. Lipids, 2016, 51(2): 263-270,Yamashita R, Tabata Y, Iga E, et al.
· Highly sensitive myosin phosphorylation analysis in the renal afferent arteriole[J]. Journal of Smooth Muscle Research, 2016, 52(0): 45-55,Takeya K.
· Functional dissection of the CroRS two-component system required for resistance to cell wall stressors in Enterococcus faecalis[J]. Journal of bacteriology, 2016, 198(8): 1326-1336,Kellogg S L, Kristich C J.
· Regulation of mitogen-activated protein kinase by protein kinase C and mitogen-activated protein kinase phosphatase-1 in vascular smooth muscle[J]. American Journal of Physiology-Cell Physiology, 2016, 310(11): C921-C930,Trappanese D M, Sivilich S, Ets H K, et al.
· ModProt: a database for integrating laboratory and literature data about protein post-translational modifications[J]. Journal of Electrophoresis, 2016, 60(1): 1-4,Kimura Y, Toda T, Hirano H.
· The C-ETS2-TFEB Axis Promotes Neuron Survival under Oxidative Stress by Regulating Lysosome Activity[J]. Oxidative medicine and cellular longevity, 2016,Ma S, Fang Z, Luo W, et al.
· Essential role of the PSI–LHCII supercomplex in photosystem acclimation to light and/or heat conditions by state transitions[J]. Photosynthesis Research, 2016: 1-10,Marutani Y, Yamauchi Y, Higashiyama M, et al.
· Identification of a redox-modulatory interaction between selenoprotein W and 14-3-3 protein[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(1): 10-18,Jeon Y H, Ko K Y, Lee J H, et al.
· Effects of hydrogen sulfide on the heme coordination structure and catalytic activity of the globin-coupled oxygen sensor AfGcHK[J]. BioMetals, 2016, 29(4): 715-729,Fojtikova V, Bartosova M, Man P, et al.
· Identification and functional analysis of phosphorylation in Newcastle disease virus phosphoprotein[J]. Archives of virology, 2016: 1-14,Qiu X, Zhan Y, Meng C, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports, 2016, 6: 54-62,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· Profiling DNA damage-induced phosphorylation in budding yeast reveals diverse signaling networks[J]. Proceedings of the National Academy of Sciences, 2016: 201602827,Zhou C, Elia A E H, Naylor M L, et al.
· Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system[J]. Nucleic Acids Research, 2016: gkw642,Brosse A, Korobeinikova A, Gottesman S, et al.
· Evolution of ZnII–Macrocyclic Polyamines to Biological Probes and Supramolecular Assembly[J]. Macrocyclic and Supramolecular Chemistry: How Izatt-Christensen Award Winners Shaped the Field, 2016: 415,Kimura E, Koike T, Aoki S.
· Phosphopeptide Enrichment Using Various Magnetic Nanocomposites: An Overview[J]. Phospho-Proteomics: Methods and Protocols, 2016: 193-209,Batalha Í L, Roque A C A.
· In vivo phosphorylation of a peptide tag for protein purification[J]. Biotechnology letters, 2016, 38(5): 767-772,Goux M, Fateh A, Defontaine A, et al.
· Regulation of cell reversal frequency in Myxococcus xanthus requires the balanced activity of CheY‐like domains in FrzE and FrzZ[J]. Molecular microbiology, 2016,Kaimer C, Zusman D R.
· Elevation of cortical serotonin transporter activity upon peripheral immune challenge is regulated independently of p38 mitogen‐activated protein kinase activation and transporter phosphorylation[J]. Journal of neurochemistry, 2016, 137(3): 423-435,Schwamborn R, Brown E, Haase J.
· The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression[J]. Molecular cell, 2016, 62(4): 532-545,Ewald J C, Kuehne A, Zamboni N, et al.
· Two Degradation Pathways of the p35 Cdk5 (Cyclin-dependent Kinase) Activation Subunit, Dependent and Independent of Ubiquitination[J]. Journal of Biological Chemistry, 2016, 291(9): 4649-4657,Takasugi T, Minegishi S, Asada A, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports. 2016,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· a high‐affinity LCO‐binding protein of Medicago truncatula, interacts with LYK3, a key symbiotic receptor[J]. FEBS letters, 2016, 590(10): 1477-1487,Fliegmann J, Jauneau A, Pichereaux C, et al. LYR3,
· Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability[J]. Molecular Cell, 2016,Spies J, Waizenegger A, Barton O, et al.
· PhostagTM-gel retardation and in situ thylakoid kinase assay for determination of chloroplast protein phosphorylation targets[J]. Endocytobiosis and Cell Research, 2016, 27(2): 62-70,Dytyuk Y, Flügge F, Czarnecki O, et al.
· Luteinizing Hormone Causes Phosphorylation and Activation of the cGMP Phosphodiesterase PDE5 in Rat Ovarian Follicles, Contributing, Together with PDE1 Activity, to the Resumption of Meiosis[J]. Biology of reproduction, 2016: biolreprod. 115.135897,Egbert J R, Uliasz T F, Shuhaibar L C, et al.
· Newby, AC, & Bond, M.(2016). The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of Molecular and Cellular Cardiology, 2016, 90: 1-10,Kimura-Wozniak T, Duggirala A, Smith M C, et al. G.
· Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels[J]. The FEBS Journal, 2016, 283(15): 2911-2928,Toume M, Tani M.
· Regulation of CsrB/C sRNA decay by EIIAGlc of the phosphoenolpyruvate: carbohydrate phosphotransferase system[J]. Molecular microbiology, 2016, 99(4): 627-639,Leng Y, Vakulskas C A, Zere T R, et al.
· The Late S-Phase Transcription Factor Hcm1 Is Regulated through Phosphorylation by the Cell Wall Integrity Checkpoint[J]. Molecular and cellular biology, 2016: MCB. 00952-15,Negishi T, Veis J, Hollenstein D, et al.
· Validation of chemical compound library screening for transcriptional co‐activator with PDZ‐binding motif inhibitors using GFP‐fused transcriptional co‐activator with PDZ‐binding motif[J]. Cancer science, 2016, 107(6): 791-802,Nagashima S, Maruyama J, Kawano S, et al.
· ULK1/2 Constitute a Bifurcate Node Controlling Glucose Metabolic Fluxes in Addition to Autophagy[J]. Molecular cell, 2016, 62(3): 359-370,Li T Y, Sun Y, Liang Y, et al.
· Spatiotemporal dynamics of Oct4 protein localization during preimplantation development in mice[J]. Reproduction, 2016: REP-16-0277,Fukuda A, Mitani A, Miyashita T, et al.
· The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress[J]. Parasites & Vectors, 2016, 9(1): 1,Pastor-Fernández I, Regidor-Cerrillo J, Álvarez-García G, et al.
· Interaction Analysis of a Two-Component System Using Nanodiscs[J]. PloS one, 2016, 11(2): e0149187,Hörnschemeyer P, Liss V, Heermann R, et al.
· Constitutive Activation of PINK1 Protein Leads to Proteasome-mediated and Non-apoptotic Cell Death Independently of Mitochondrial Autophagy[J]. Journal of Biological Chemistry, 2016, 291(31): 16162-16174,Akabane S, Matsuzaki K, Yamashita S, et al.
· p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261[J]. Molecular and cellular biology, 2016, 36(10): 1540-1554,Beenstock J, Melamed D, Mooshayef N, et al.
· Lysophosphatidylcholine acyltransferase 1 protects against cytotoxicity induced by polyunsaturated fatty acids[J]. The FASEB Journal, 2016, 30(5): 2027-2039,Akagi S, Kono N, Ariyama H, et al.
· Characterization of a herpes simplex virus 1 (HSV-1) chimera in which the Us3 protein kinase gene is replaced with the HSV-2 Us3 gene[J]. Journal of virology, 2016, 90(1): 457-473,Shindo K, Kato A, Koyanagi N, et al.
· Generation of phospho‐ubiquitin variants by orthogonal translation reveals codon skipping[J]. FEBS letters, 2016, 590(10): 1530-1542,George S, Aguirre J D, Spratt D E, et al.
· Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris[J]. PLoS Genet, 2016, 12(3): e1005922,Ma P, Mori T, Zhao C, et al.
· Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis[J]. FEMS Yeast Research, 2016, 16(6): fow060,Pérez J, Arcones I, Gómez A, et al.
· Alteration of Antiviral Signalling by Single Nucleotide Polymorphisms (SNPs) of Mitochondrial Antiviral Signalling Protein (MAVS)[J]. PloS one, 2016, 11(3): e0151173,Xing F, Matsumiya T, Hayakari R, et al.
· Arm-in-arm response regulator dimers promote intermolecular signal transduction[J]. Journal of bacteriology, 2016, 198(8): 1218-1229,Baker A W, Satyshur K A, Morales N M, et al.
· The lsh/ddm1 homolog mus-30 is required for genome stability, but not for dna methylation in neurospora crassa[J]. PLoS Genet, 2016, 12(1): e1005790,Basenko E Y, Kamei M, Ji L, et al.
· Fine tuning chloroplast movements through physical interactions between phototropins[J]. Journal of Experimental Botany, 2016: erw265,Sztatelman O, Łabuz J, Hermanowicz P, et al.
· Characterization of the Neospora caninum NcROP40 and NcROP2Fam-1 rhoptry proteins during the tachyzoite lytic cycle[J]. Parasitology, 2016, 143(01): 97-113,Pastor-Fernandez I, Regidor-Cerrillo J, Jimenez-Ruiz E, et al.
· Transcriptional Profile during Deoxycholate-Induced Sporulation in a Clostridium perfringens Isolate Causing Foodborne Illness[J]. Applied and environmental microbiology, 2016, 82(10): 2929-2942,Yasugi M, Okuzaki D, Kuwana R, et al.
· Timely Closure of the Prospore Membrane Requires SPS1 and SPO77 in Saccharomyces cerevisiae[J]. Genetics, 2016: genetics. 115.183939,Paulissen S M, Slubowski C J, Roesner J M, et al.
· DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach[J]. Nucleic Acids Research, 2016: gkw626,Wu K Z L, Wang G N, Fitzgerald J, et al.
· OVATE Family Protein 8 Positively Mediates Brassinosteroid Signaling through Interacting with the GSK3-like Kinase in Rice[J]. PLoS Genet, 2016, 12(6): e1006118,Yang C, Shen W, He Y, et al.
· Epithelial Sel1L is required for the maintenance of intestinal homeostasis[J]. Molecular biology of the cell, 2016, 27(3): 483-490, Sun S, Lourie R, Cohen S B, et al.
· Effect of Sodium Dodecyl Sulfate Concentration on Supramolecular Gel Electrophoresis[J]. ChemNanoMat, 2016,Tazawa S, Kobayashi K, Yamanaka M.
· Intergenic VNTR Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and immunity, 2016: IAI. 00258-16,Zhu L, Olsen R J, Horstmann N, et al.
· Ajuba Phosphorylation by CDK1 Promotes Cell Proliferation and Tumorigenesis[J]. Journal of Biological Chemistry, 2016: jbc. M116. 722751,Chen X, Stauffer S, Chen Y, et al.
· Editorial: International Plant Proteomics Organization (INPPO) World Congress 2014[J]. Frontiers in Plant Science, 2016, 7,Heazlewood J L, Jorrín-Novo J V, Agrawal G K, et al.
· Phosphoinositide kinase signaling controls ER-PM cross-talk[J]. Molecular biology of the cell, 2016, 27(7): 1170-1180,Omnus D J, Manford A G, Bader J M, et al.
· A multiple covalent crosslinked soft hydrogel for bioseparation[J]. Chemical Communications, 2016, 52(15): 3247-3250,Liu Z, Fan L, Xiao H, et al.
· Advances in crop proteomics: PTMs of proteins under abiotic stress[J]. Proteomics, 2016, 16(5): 847-865,Wu X, Gong F, Cao D, et al.
· Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae[J]. Molecular cell, 2016, 62(4): 546-557,Zhao G, Chen Y, Carey L, et al.
· Carbon Monoxide Gas Is Not Inert, but Global, in Its Consequences for Bacterial Gene Expression, Iron Acquisition, and Antibiotic Resistance[J]. Antioxidants & redox signaling, 2016,Wareham L K, Begg R, Jesse H E, et al.
· Two-layer regulation of PAQR3 on ATG14-linked class III PtdIns3K activation upon glucose starvation[J]. Autophagy, 2016: 1-2,Xu D, Wang Z, Chen Y.
· Regulation of sphingolipid biosynthesis by the morphogenesis checkpoint kinase Swe1[J]. Journal of Biological Chemistry, 2016, 291(5): 2524-2534,Chauhan N, Han G, Somashekarappa N, et al.
· PAX5 tyrosine phosphorylation by SYK co-operatively functions with its serine phosphorylation to cancel the PAX5-dependent repression of BLIMP1: A mechanism for antigen-triggered plasma cell differentiation[J]. Biochemical and biophysical research communications, 2016, 475(2): 176-181,Inagaki Y, Hayakawa F, Hirano D, et al.
· A Combined Computational and Genetic Approach Uncovers Network Interactions of the Cyanobacterial Circadian Clock[J]. Journal of Bacteriology, 2016: JB. 00235-16,Boyd J S, Cheng R R, Paddock M L, et al.
· HuR mediates motility of human bone marrow-derived mesenchymal stem cells triggered by sphingosine 1-phosphate in liver fibrosis[J]. Journal of Molecular Medicine, 2016: 1-14,Chang N, Ge J, Xiu L, et al.
· Combined replacement effects of human modified β-hexosaminidase B and GM2 activator protein on GM2 gangliosidoses fibroblasts[J]. Biochemistry and Biophysics Reports, 2016,Kitakaze K, Tasaki C, Tajima Y, et al.
· Roseotoxin B Improves Allergic Contact Dermatitis through a Unique Anti-inflammatory Mechanism Involving Excessive Activation of Autophagy in Activated T-Lymphocytes[J]. Journal of Investigative Dermatology, 2016,Wang X, Hu C, Wu X, et al.
References on Phos-tag™ Chemistry
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule, Rapid Communications of Mass Spectrometry, 17, 2075-2081 (2003), H. Takeda, A. Kawasaki, M. Takahashi, A. Yamada, and T. Koike
Recognition of phosphate monoester dianion by an alkoxide-bridged dinuclear zinc (II) complex, Dalton Transactions, 1189-1193 (2004), E. Kinoshita, M. Takahashi, H. Takeda, M. Shiro, and T. Koike
Quantitative analysis of lysophosphatidic acid by time-of-flight mass spectrometry using a phosphate capture molecule, Journal of Lipid Research, 45, 2145-2150 (2004), T. Tanaka, H. Tsutsui, K. Hirano, T. Koike, A. Tokumura, and K. Satouchi
Production of 1,2-Didocosahexaenoyl Phosphatidylcholine by Bonito Muscle Lysophosphatidylcholine/Transacylase, Journal of Biochemistry,136, 477-483 (2004), K. Hirano, H. Matsui, T. Tanaka, F. Matsuura, K. Satouchi, and T. Koike
Novel immobilized zinc(II) affinity chromatography for phosphopeptides and phosphorylated proteins, Journal of Separation Science, 28, 155-162 (2005), E. Kinoshita, A. Yamada, H. Takeda, E. Kinoshita-Kikuta, and T. Koike
Detection and Quantification of On-Chip Phosphorylated Peptides by Surface Plasmon Resonance Imaging Techniques Using a Phosphate Capture Molecule, Analytical Chemistry, 77, 3979-3985 (2005), K. Inamori, M. Kyo, Y. Nishiya, Y. Inoue, T. Sonoda, E. Kinoshita, T. Koike, and Y. Katayama
Phosphate-binding tag: A new tool to visualize phosphorylated proteins, Molecular & Cellular Proteomics, 5, 749-757 (2006), E. Kinoshita, E. Kinoshita-Kikuta, K. Takiyama, and T. Koike
Enrichment of phosphorylated proteins from cell lysate using phosphate-affinity chromatography at physiological pH, Proteomics, 6, 5088-5095 (2006), E. Kinoshita-Kikuta, E. Kinoshita, A. Yamada, M. Endo, and T. Koike
Separation of a phosphorylated histidine protein using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 360, 160-162 (2007), S. Yamada, H. Nakamura, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and Y. Shiro
Label-free kinase profiling using phosphate-affinity polyacrylamide gel electrophresis, Molecular & Cellular Proteomics, 6, 356-366 (2007), E. Kinoshita-Kikuta, Y. Aoki, E. Kinoshita, and T. Koike
A SNP genotyping method using phosphate-affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 361, 294-298 (2007), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike (The phosphate group at DNA-terminal is efficiently captured by Zn2+.Phos-tag.)
Identification on Membrane and Characterization of Phosphoproteins Using an Alkoxide-Bridged Dinuclear Metal Complex as a Phosphate-Binding Tag Molecule, Journal of Biomolecular Techniques, 18, 278-286 (2007), T. Nakanishi, E. Ando, M. Furuta, E. Kinoshita, E. Kikuta-Kinoshita, T. Koike, S. Tsunasawa, and O. Nishimura
A mobility shift detection method for DNA methylation analysis using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 378, 102-104 (2008), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
Separation of phosphoprotein isotypes having the same number of phosphate groups using phosphate- affinity SDS-PAGE, Proteomics, 8, 2994-3003 (2008), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, S. Yamada, H. Nakamura, Y. Shiro, Y. Aoki, K. Okita, and T. Koike
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway, Nature Structural & Molecular Biology, 15, 1138-1146 (2008), M. Ishiai, H. Kitao, A. Smogorzewska, J. Tomida, A. Kinomura, E. Uchida, A. Saberi, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, S. Tashiro, S. J. Elledge, and M. Takata
Two-dimensional phosphate affinity gel electrophoresis for the analysis of phosphoprotein isotypes , Electrophoresis, 30, 550-559 (2009), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, Y. Aoki, S. Ohie, Y. Mouri, and T. Koike
Formation of lysophosphatidic acid, a wound-healing lipid, during digestion of cabbage leaves, Bioscience, Biotechnology, and Biochemistry,73, 1293-300 (2009), T. Tanaka, G. Horiuchi, M. Matsuoka, K. Hirano, A. Tokumura, T. Koike, and K. Satouchi
A Phos-tag-based fluorescence resonance energy transfer system for the analysis of the dephosphorylation of phosphopeptides, Analytical Biochemistry, 388, 235-241, (2009), K. Takiyama, E. Kinoshita, E. Kinoshita-Kikuta, Y. Fujioka, Y. Kubo, and T. Koike
Phos-tag beads as an immunoblotting enhancer for selective detection of phosphoproteins in cell lysates, Analytical Biochemistry, 389, 83-85, (2009), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
Mobility shift detection of phosphorylation on large proteins using a Phos-tag SDS-PAGE gel strengthened with agarose, Proteomics, 9, 4098- 4101 (2009), E. Kinoshita, E. Kinoshita-Kikuta, H. Ujihara, and T. Koike
Separation and detection of large phosphoproteins using Phos-tag SDS-PAGE, Nature Protocols, 4, 1513-1521 (2009), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike
A clean-up technology for the simultaneous determination of lysophosphatidic acid and sphingosine-1-phosphate by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a phosphate-capture molecule, Phos-tag, Rapid Communications in Mass Spectrometry, 24, 1075-1084 (2010), J. Morishige, M. Urikura, H. Takagi, K. Hirano, T. Koike, T. Tanaka, and K. Satouchi
Genotyping and mapping assay of single-nucleotide polymorphisms in CYP3A5 using DNA-binding zinc(II) complexes, Clinical Biochemistry, 43, 302-306 (2010), E. Kinoshita, E. Kinoshita-Kikuta, H. Nakashima, and T. Koike
The DNA-binding activity of mouse DNA methyltransferase 1 is ragulated phosphorylation with casein kinase 1σ/ε, Biochemical Journal, 427, 489-497 (2010), Y. Sugiyama, N. Hatano, N. Sueyoshi, I. Suetake, S. Tajima, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and I. Kameshita
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 308-93563 | Phos-tag™ Agarose Phos-tag 琼脂糖 |
3 mL | – |
KANEKA KanCapA™ 新标准 抗体纯化树脂
- 产品特性
- 相关资料
- Q&A
- 参考文献
新标准 抗体纯化树脂
KANEKA KanCapA™

本产品为 Protein A 亲和层析树脂。用于从活体样本及细胞培养上清纯化免疫球蛋白及包含 Fc 段的重组抗体。
本产品由高交联纤维素微珠及新研发的耐碱 Protein A 配体构成,可用于 IgG 的亲和纯化。
◆特点
• 耐碱
• 良好洗脱条件
• 高抗体结合能力
• 可循环使用
• 便于扩增
◆关于改良 Protein A

KanCapA™ 采用的不是天然 Protein A,而是使用变异型C域5聚物的新型 Protein A。
天然 Protein A的C域结合于 Fc 段,也结合于 Fab 段,其中与VH3亚家族抗体的反应性较高。该类型的解离需要弱酸条件下洗脱。
为改善该问题,使用分子设计系统重新设计分子,研发了保持耐碱性、改善 Fab 段的非特异性结合及洗脱曲线的 Protein A。
本产品采用的 Protein A
|
1. C5 聚体蛋白质 |
 |
良好pH条件下清晰地洗脱抗体 |
|
2. 较高的耐碱性 |
||
|
3. 减少 Fab 段的非特异性结合 |
◆产品特性
KANEKA KanCapA™ 产品特性
|
矩阵 |
高交联纤维 |
※ 1) 平均粒度为累积体积分布的中间粒度 2) 5%动态结合容量为前端分析测定(停留时间3分钟) 3) 可在强碱条件下清洗,但请避免长时间的清洗 4) 请勿冷冻 |
|
平均粒度1) |
65-85 μm |
|
|
配体 |
Protein A,重组(抗强碱) |
|
|
结合 |
化学结合(还原胺化) |
|
|
动态结合容量2) |
≧35 mg 人多克隆 IgG/mL载体 |
|
|
化学稳定性3) |
使用亲和层析通用可溶性溶媒时较稳定 |
|
|
pH 范围3) |
pH 2-13 |
|
|
静止清洗条件3) |
0.1-0.5 mol/L氢氧化钠 6-8 mol/L尿素、6 mol/L盐酸胍 |
|
|
流量 |
最大 500cm/h(层高:20-25 cm) |
|
|
停留时间 |
≧3分钟(建议:4-6分钟) |
|
|
保存条件4) |
20%乙醇浑浊液,1~10℃ |
◆应用
1、Fab段 纯化应用
• 利用 KANEKA KanCapA™ 的特点,可从经木瓜蛋白酶处理后的 IgG 溶液分馏 Fc 段及 Fab 段。

2、IgG 洗脱曲线(pH3.5 & pH3.0)
• 洗脱范围已优化,可清晰地分馏提纯。

3、通过强碱清洗进行树脂循环
• 进行 300 次循环处理后(0.1M 氢氧化钠溶液 15 分钟),IgG 的结合容量的减少率仍低于20%,表明其循环性能较高。


|
使用 0.1 M 氢氧化钠溶液进行循环处理后的 IgG 结合活性曲线数据 |
循环处理后(0.1 M 氢氧化钠溶液 15 分钟)的IgG回收率及 Protein A 配体的渗滤量的曲线数据 |
4、动态结合容量 曲线数据
• 在4-6分钟停留时间过程中,DBC 较高。
※ 动态结合容量表示纯化样本在流动的状态下,回收程度的数值。动态结合容量较高的载体的流速较快,可回收大量目标分子,因此可以认为在短时间内有效纯化目标分子的优质载体。

KANEKA KanCapA™的Dynamic Binding Capacity (DBC; 动态结合容量) 曲线数据
5、不同大小的树脂填充装柱的压力/流速绘图数据

填充依赖压力的计算方法是从整体压力减去系统压力。
6、KANEKA KanCapA™ 于各种大小的柱的填充效果
• KANEKA KanCapA™ 适用于各种大小的柱。
|
充填方法 |
L. D.(cm) |
高度(mm) |
培养皿/m |
非对称性 |
|
流动填充 |
4.4 |
200 |
5710 |
1.14 |
|
轴向力填充 |
7 |
199 |
6063 |
1.06 |
|
流动填充 |
10 |
201 |
5276 |
1.18 |
|
流动填充 |
30 |
200 |
5208 |
1.17 |
|
轴向力填充 |
45 |
233 |
4712 |
1.09 |
以下是脉冲注入数据:
可溶性溶媒:1(v/v)% 丙酮
注入量:1% 柱容量
流速:60 cm/h


参考文献
使用Kaneka KanCapA纯化单克隆抗体
使用该Kaneka KanCapATM从CHO细胞培养上清中纯化单克隆抗体
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 114-01071 | KANEKA KanCapA™ | 2mL (net 1mL) | for Genetic Research |
| 110-01073 | KANEKA KanCapA™ | 10mL (net 5mL) | for Genetic Research |
| 118-01074 | KANEKA KanCapA™ | 50mL (net 25mL) | for Genetic Research |
MagCapture™ 外泌体提取试剂盒 PS Ver.2 提高回收率和纯化纯度的外泌体提取/纯化试剂盒
- 产品特性
- 相关资料
- Q&A
- 参考文献
提高回收率和纯化纯度的外泌体提取/纯化试剂盒![]()
MagCapture™ 外泌体提取试剂盒 PS Ver.2
本试剂盒是提取/纯化外泌体的试剂盒,比起第一代产品(293-77601 MagCapture™ 外泌体提取试剂盒PS),提高了外泌体的回收率和纯度,并缩短了实验操作的时间。使用本试剂盒纯化的外泌体可以直接用于细胞摄取实验。
◆特点
● 提高了外泌体的回收率和纯度
● 缩短亲和反应的时间(3 h→1 h)
● 降低纯化外泌体的细胞毒性
◆数据
■ 比较外泌体的回收率
|
纳米级粒子数检测 |
ELISA EV表面标记物的比较 |
|
|
|


Western blotting EV标记的比较

在间充质干细胞(MSC)的培养上清中,使用第一代产品(图中简写为Ver.1)和本试剂盒(图中简写为Ver.2)提取和纯化含外泌体的细胞外囊泡(图中简写为EV)。然后,对所得样品进行纳米级粒子数检测以及使用PS MagCapture™ 外泌体 ELISA Kit(链霉亲和素HRP)(产品编号:298-80601)和WB比较EV标记物。
Ver.2与Ver.1相比,回收率有所提高。
■ 比较纯化外泌体的细胞毒性

使用Ver.1和Ver.2提取和纯化COLO201细胞培养上清中含外泌体的细胞外囊泡。然后仅将elution buffer或纯化外泌体加入到人正常成纤维细胞中,观察经过48 h后的细胞形态变化。
在Ver.1中,加入外泌体和elution buffer后48 h出现了细胞死亡,而Ver.2中未观察到明显的细胞毒性。证明Ver.2中elution buffer的细胞毒性已经降低了。
■ 比较不同亲和反应时间里外泌体纯化的回收率

在COLO201细胞培养上清中,为Ver.2设定4个亲和反应时间条件,提取和纯化含外泌体的细胞外囊泡(EV)。然后,获得的样品使用PS MagCapture™ 外泌体 ELISA 试剂盒(链霉亲和素HRP)(产品编号:298-80601)比较EV表面标记物。
在Ver.2中,确认了1 h的亲和反应可以充分提取到培养上清中的细胞外囊泡。
※ 本页面产品仅供研究用,研究以外不可使用。
| 产品编号 | 产品名称 | 产品规格 | 产品等级 |
| 294-84101 | MagCapture™ Exosome Isolation Kit PS Ver.2 MagCapture™ 外泌体提取试剂盒 PS Ver.2 |
2次用 | 基因研究用 |
| 290-84103 | MagCapture™ Exosome Isolation Kit PS Ver.2 MagCapture™ 外泌体提取试剂盒 PS Ver.2 |
10次用 | 基因研究用 |
NucleoFast 96 PCR 核酸纯化板 NucleoFast 96 PCR Plates
NucleoFast 96 PCR 核酸纯化板 NucleoFast 96 PCR Plates
NucleoFast 96 PCR 核酸纯化板 NucleoFast 96 PCR Plates
Protino 纯化板 Protino Purification Plate
Protino 纯化板 Protino Purification Plate
慢病毒纯化酶试剂盒Takara
上海金畔生物科技有限公司代理Takara酶试剂盒全线产品,欢迎访问官网了解更多产品信息。
| 慢病毒纯化 | ||||||
| 品牌 | Code No. | 产品名称 | 包装量 | 价格(元) | 说明书 | 数量 |
| Clontech | 631246 | Collection Rack L | Each | ¥1,469 | |
  |
| Clontech | 631233 | Lenti-X Maxi Purification Kit | 2 Preps | ¥3,287 | |
|
| Clontech | 631234 | Lenti-X Maxi Purification Kit | 5 Preps | ¥5,474 | |
|
| Clontech | 631245 | Lenti-X Maxi Purification Kit (with Rack) | Each | ¥6,864 | |
|
收藏产品 加入购物车
.png)
.png)
.png)
.png)
.png)
.png)
.png)

页面更新:2020-02-25 11:41:30
Collagenase, Purified 胶原酶,纯化 品牌:Wako
|
品牌:Wako
CAS No.:9001-12-1 储存条件:2-10℃ 纯度:– |
| 产品编号
(生产商编号) |
等级 | 规格 | 运输包装 | 零售价(RMB) | 库存情况 | 参考值 |
|
031-24073 |
– | 25000 units | – | 咨询 | – | – |
* 干冰运输、大包装及大批量的产品需酌情添加运输费用
* 零售价、促销产品折扣、运输费用、库存情况、产品及包装规格可能因各种原因有所变动,恕不另行通知,确切详情请联系上海金畔生物科技有限公司。
产品描述相关资料下载相关产品浏览记录 请联系客服
Collagenase, Purified 胶原酶,纯化 品牌:Wako
|
品牌:Wako
CAS No.:9001-12-1 储存条件:2-10℃ 纯度:– |
| 产品编号
(生产商编号) |
等级 | 规格 | 运输包装 | 零售价(RMB) | 库存情况 | 参考值 |
|
035-24071 |
– | 5000 units | – |
|
– | – |

* 干冰运输、大包装及大批量的产品需酌情添加运输费用
* 零售价、促销产品折扣、运输费用、库存情况、产品及包装规格可能因各种原因有所变动,恕不另行通知,确切详情请联系上海金畔生物科技有限公司。
产品描述相关资料下载相关产品浏览记录 请联系客服