
上海金畔生物科技有限公司代理AAT Bioquest荧光染料全线产品,欢迎访问AAT Bioquest荧光染料官网了解更多信息。
Screen Quest CHO-Gqi嵌合体细胞系
| 货号 | 38101 | 存储条件 | 在零下80度以下保存 |
| 规格 | Each | 价格 | 0 |
| Ex (nm) | Em (nm) | ||
| 分子量 | 溶剂 | ||
| 产品详细介绍 | |||
简要概述
产品基本信息
货号:38101
产品名称:Screen Quest CHO-Gqi嵌合体细胞系
规格:Each
适用仪器
| 其他仪器 |
| FDSS, FLIPR, ViewLux, NOVOStar, ArrayScan, FlexStation, IN Cell Analyzer |
产品介绍
Screen Quest 细胞系是一系列细胞,已成功用于药物发现和筛选环境中,用于研究通常不通过细胞内钙偶联的G蛋白偶联受体(GPCR)。它已与FLIPR,FDSS系统有效结合,与许多受体的非Gq偶联成员结合使用,例如趋化因子,5-羟色胺,谷氨酸,多巴胺,阿片样物质,血管加压素以及α和β-肾上腺素能受体家族。超过60%的已知G蛋白偶联受体通过Gq以外的途径发出信号,从而导致细胞内钙增加,并且随着基因组学揭示出更多的GPCR靶标,这种趋势持续增加。Screen Quest 细胞系用于研究通常不通过细胞内钙偶联的GPCR。Screen Quest 细胞系基于一系列G蛋白嵌合体,包括混杂的G蛋白Gα16。嵌合体由Gq蛋白复合物的α亚基组成,该复合物的5个羧基末端氨基酸已被其他G蛋白之一(Gαs,Gαi,Gαo或Gαz)取代。这些氨基酸负责受体与其G蛋白的偶联。这些嵌合体与通常通过cAMP途径起作用的特定非Gq偶联受体的共表达可能会导致受体刺激后细胞内钙信号的产生。Screen Quest CHO-Gqi细胞系是被嵌合Gqi alpha亚基蛋白稳定转染的CHO-K1细胞。当用作转染表达Gi偶联受体的宿主细胞时,细胞中组成型表达的Gqi蛋白可使通常通过cAMP途径起作用的转染受体与Gq信号转导和动员的细胞内钙偶联。可以使用钙敏感染料(例如Calbryte 520 AM,Cal-520 AM,Fluo-8 AM或Fluo-4 AM)和无需洗涤的钙试剂盒检测特定配体对这些非Gq偶联受体的激活。金畔生物是AAT Bioquest的中国代理商,为您提供最优质的Screen Quest CHO-Gqi嵌合体细胞系。
图示
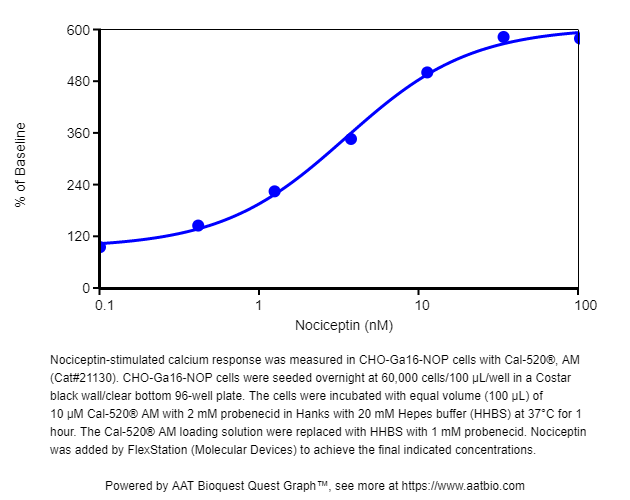 图1.用Cal-520®,AM(Cat#21130)在CHO-Ga16-NOP细胞中测量了伤害感受肽刺激的钙反应。将CHO-Ga16-NOP细胞以60,000个细胞/ 100 µL /孔在Costar黑色96孔板中过夜接种。在含有20 mM Hepes缓冲液(HHBS)的Hanks中,将等体积(100μL)的10μMCal-520®AM和2 mM丙磺舒在37°C下孵育1小时。用具有1 mM丙磺舒的HHBS代替Cal-520®AM上样溶液。通过FlexStation(分子设备)添加了Nociceptin,以达到最终指示的浓度。 |
参考文献
A cardiac mitochondrial cAMP signaling pathway regulates calcium accumulation, permeability transition and cell death
Authors: Wang Z, Liu D, Varin A, Nicolas V, Courilleau D, Mateo P, Caubere C, Rouet P, Gomez AM, V and ecasteele G, Fischmeister R, Brenner C.
Journal: Cell Death Dis (2016): e2198
Activation of P2X7 and P2Y11 purinergic receptors inhibits migration and normalizes tumor-derived endothelial cells via cAMP signaling
Authors: Avanzato, D and Genova, T and Pla, A Fiorio and Bernardini, M and Bianco, S and Bussolati, B and Mancardi, D and Giraudo, E and Maione, F and Cassoni, P and others
Journal: Scientific Reports (2016)
Changes in the Arabidopsis thaliana Proteome Implicate cAMP in Biotic and Abiotic Stress Responses and Changes in Energy Metabolism
Authors: Alqurashi M, Gehring C, Marondedze C.
Journal: Int J Mol Sci (2016): 852
Odor-induced cAMP production in Drosophila melanogaster olfactory sensory neurons
Authors: Miazzi F, Hansson BS, Wicher D.
Journal: J Exp Biol (2016): 1798
Role of the cAMP Pathway in Glucose and Lipid Metabolism
Authors: Ravnskjaer K, Madiraju A, Montminy M.
Journal: Handb Exp Pharmacol (2016): 29
The pleiotropic role of exchange protein directly activated by cAMP 1 (EPAC1) in cancer: implications for therapeutic intervention
Authors: Almahariq M, Mei FC, Cheng X.
Journal: Acta Biochim Biophys Sin (Shanghai) (2016): 75
cAMP-Induced Histones H3 Dephosphorylation Is Independent of PKA and MAP Kinase Activations and Correlates With mTOR Inactivation
Authors: Rodriguez P, Rojas J.
Journal: J Cell Biochem (2016): 741
A cAMP Biosensor-Based High-Throughput Screening Assay for Identification of Gs-Coupled GPCR Ligands and Phosphodiesterase Inhibitors
Authors: Vedel L, Brauner-Osborne H, Mathiesen JM.
Journal: J Biomol Screen (2015): 849
Cardiac Hypertrophy Is Inhibited by a Local Pool of cAMP Regulated by Phosphodiesterase 2
Authors: Zoccarato A, Surdo NC, Aronsen JM, Fields LA, Mancuso L, Dodoni G, Stangherlin A, Livie C, Jiang H, Sin YY, Gesellchen F, Terrin A, Baillie GS, Nicklin SA, Graham D, Szabo-Fresnais N, Krall J, V and eput F, Movsesian M, Furlan L, Corsetti V, Hamilton G, Lefkimmiatis K, Sjaastad I, Zaccolo M.
Journal: Circ Res (2015): 707
Cardiac cAMP: production, hydrolysis, modulation and detection
Authors: Boularan C, Gales C.
Journal: Front Pharmacol (2015): 203